

서 론
옥수수(Zea mays L.)는 밀(Triticum aesticum L.), 벼(Oryza sativa L.)와 함께 세계 3대 식량작물 중 하나로 전 세계적으로 오랜 재배역사를 가지고 있으며 식용, 사료용, 공업용 및 바이오에너지용 등 다양한 용도로 이용되고 있다(Heo et al., 2017). 단위면적당 건물생산성과 영양가, 식미 등 기호성이 좋아 국내에서는 조선시대 이후 쌀이나 보리 등 주식작물 대용으로 널리 재배되어 온 대표적인 작물이며, 현재 국내의 풋옥수수의 재배면적은 15,000 ha로 매년 증가하는 추세이다(Shim et al., 2019). 옥수수는 주로 풋찰옥수수 상태로 간식으로 이용되기 때문에 재배기간이 다른 작물 대비 85~110일 정도로 짧아 2모작이나 2기작 재배가 가능하며(Park et al., 2016), 간식용으로 재배되는 풋옥수수는 과거 경립종도 이용되었지만, 현재는 찰옥수수, 단옥수수, 초당옥수수가 주로 재배되고 있다(Yang et al., 2007). 찰옥수수를 노지에서 재배할 경우 조수익이 113~153만원/10a 정도로 관광지 및 브랜드화가 정착된 지역에서는 더 높은 소득이 가능하며, 조기 출하하는 경남의 시설재배에서 조수익이 300만원에 이르는 등 수익성이 비교적 높은 작물 중 하나이다(Park et al., 2016; Jeon & Cho, 2019).
특히, 찰옥수수 일반재배의 경우 수확기인 여름철에 홍수출하로 인하여 가격폭락 문제가 매년 발생하고 있으며, 이러한 문제를 피하기 위하여 생육 초기에 저온 피해가 없는 범위 내에서 가급적 일찍 파종하는 것이 수량을 높일 수 있다(Heo et al., 2017; Jeon & Cho, 2019). 사일리지용 옥수수도 4월 중순 파종하여 재배할 경우 수확기에 가을장마와 태풍을 피하기 위해 조기 수확한 미숙상태의 옥수수를 가지고 제조하여 수량 및 품질저하 등의 문제점이 야기되고 있어(Kim et al., 1996), 식용옥수수의 문제만이라고는 할 수 없다. 이러한 조건하에서는 옥수수를 가능한 일찍 파종해서 수확기까지의 충분한 생육기간을 확보하여 주는 것이 수량의 증수 및 품질향상의 적절한 방법이라 할 수 있고, 옥수수 재배농가가 옥수수의 수량을 추가로 높일 수 있는 유일한 방법은 옥수수의 조기파종이라 지적한 바 있다(Aldrich et al., 1986 & Kim et al., 1996). 최근 기후온난화와 같은 기상이변으로 발생되는 자연재해에 의해 생물학적 및 비생물학적 환경 스트레스(biotic and abiotic stress)로 작물의 생산성이 매년 불안정한 상태이다(Seo et al., 2014). 온도는 옥수수 생육에 가장 큰 영향을 미치는 기상 요인 중 하나로 옥수수를 너무 일찍 파종하게 되면 저온, 다습한 조건이 되므로 파종에서 출아까지의 소요일수가 증가하고, 생육이 불량하게 된다(Shim et al., 2019). 남부지방에서도 3~4월에 꽃샘추위 등으로 인하여 파종 및 출현기에 저온에 노출되고 있으며, 3월에 눈이 내리거나 3~4월의 일평균기온이 10°C 이하인 날이 2018년에 24일, 2019년에 28일로 기록되는 등(ACR, 2019)의 기상이변으로 인하여 출현후 유묘기 시기에 다양한 피해가 발생하고 있는 실정이다.
저온장해에 대한 다른 작물의 연구는 보리 및 밀 등의 비닐피복을 이용한 저온장해 유도시 수잉기에 가장 민감한 반응을 보이며, 수량구성요소 중 수수와 일수립수에 영향을 미쳤다고 하였고(Koo et al., 2007), Han et al. (1986)은 온도변화에 따른 발아율 실험에서 10/5°C에서 수수×수단그라스 80%, 옥수수 43.2%, 수단그라스 24.8%, 수수 68.8%로 발아율이 감소하였다고 보고하였으며, Lee et al. (1977)은 수도에서 품종간 저온처리시 초장 신장이 정지되고, 저온 처리후 정상온도에 옮겼을 경우 2일까지 장해가 증가하다 그 이후에는 차이가 없었으며, 4엽기에 피해가 컸다고 하였다. 그리고 Kwon et al. (1979)은 수도 품종별 유묘기 저온연구에서는 저온처리시 적변고사현상이 일어나고, 처리기간의 증가 및 노엽일수록 피해가 증가하였으며, 내냉한계는 4일 미만 저온처리였다고 하였다. 또 저온처리후 초장이 증가하지 않았고, 처리완료후 일부 품종은 2일처리에서 대조구와 차이가 없었으며, 저온처리기간중 생장이 정체된 차이를 보상하지 못했다고 보고하였다. 이처럼 저온에 따른 냉해에 대한 연구는 맥류 및 수도작에 한하여 진행되어 왔으나, 옥수수의 경우 파종기에 관련된 연구(Lee, 1998; Yang et al., 2007; Shim et al., 2019)와 수확후 저온저장(Han et al., 1986; Kim et al., 2011) 등의 연구내용은 보고되고 있으나, 유묘기에 저온피해에 대한 피해양상에 대한 연구는 없는 실정이다.
따라서 본 연구는 풋찰옥수수의 유묘기 저온에 따른 생육특성 변화 및 피해양상을 알아봄으로써 남부지방에서 풋찰옥수수 안정적인 2모작(풋찰옥수수-콩) 또는 2기작 작부체계 확립을 위한 기초자료로 활용하고자 수행하였다.
재료 및 방법
공시재료
본 시험은 대표 찰옥수수 보급종인 일미찰 옥수수(cv. Ilmichal, 중만생종)(Kim et al., 2009) 조기파종 재배시 유묘기의 저온에 따른 생육특성을 분석하기 위하여, 국립식량과학원에서 2019년 생산된 종자를 보급받아 사용하였다.
처리방법
실험의 유묘의 선발은 초엽기, 제 1엽기, 제 2엽기로 생장시기를 3 수준으로 설정하여 수행하였다. 유묘재배는 다른 외부 환경 특성을 최대한 배제하기 위해, 와그너포트(1/5000)에 상토를 채운 후 1립씩 파종하였고, 저면관수하여 생육장(GC-1000, JEIO TECH)에서 항온 26°C 조건에서 출현시켜 각 생육시기별로 재배한 후 실험에 이용하였다. 실험을 수행한 경남 진주지방의 최근 10년간 기상자료를 바탕으로 하여 온도는 4수준으로 5, 3, 0, -3°C (±1°C) 설정하였고, 저온 처리기간은 3, 5, 7일로 3 수준으로 설정하여 20개체 3반복 수행하여 생육특성을 조사하였고, 각각 저온배양기(DS-53FPL-2C, DASOL SCIENSE)로 저온처리 하였다. 저온처리후 생육장에 26°C 조건으로 3일간 처리하여 회복 가능여부 및 생육특성을 조사하였다.
생육특성조사
생육특성조사는 저온처리 시작 직전과 저온처리가 끝나는 시점 및 처리 후 항온 생장상에 환원한 3일후로 3회 실시하였다. 농업과학기술조사 연구조사분석기준에 의거하여 유묘의 초장, 경태, 엽수, 엽색도(SPAD), 식생지수(NDVI), 엽록소 형광반응 측정(Fv/Fm)을 조사하였다. 엽색도(SPAD)는 Chlorophyll Meter SPAD-502 Plus (Japan)를 이용하여 전개가 완료된 상위 잎의 중간지점에서 측정하였고, 식생지수(NDVI)는 Photon Systems Instruments NDVI310 (Czech Republic)을 이용하여 엽색도와 동일한 지점에서 측정하였다. 엽록소 형광반응 측정(Fv/Fm)도 엽색도와 동일한 지점에서 수행하였으며, 엽을 20분 이상 엽챔버에 물려서 암상태를 진행하여 내부 환경을 안정시킨 후, Fluorpen FP-100 (PSI, Czech)을 사용하여 측정 후 광화학반응(OJIP) 측정한 값에서 산출하였다.
저온 피해양상 조사
생육시기별 저온처리기간 및 온도에 따른 피해양상 조사는 저온처리 직후 및 저온처리 완료 3일후 2회 조사하였고, 엽의 위조가 시작된 엽의 개수 및 엽의 위조 면적을 육안으로 조사하여 냉해가 없는 개체를 0으로 하고, 피해면적에 따라 1 (10% 이하), 2 (11~20%), 3 (21~30%), 4 (31~40%), 5 (41~50%), 6 (51~60%), 7 (61~70%), 8 (71~80%) 및 고사한 개체를 9로 하여 0~9까지의 지수로 환산하여 나타냈다.
통계처리
본 연구에서 얻어진 데이터의 통계분석은 SAS프로그램(V. 9.4, Cary, NC, USA)을 이용하여 분산분석하였고, Duncan의 다중검정법(Duncan’s multiple range test, DMRT)을 통해 5% 유의수준에서 처리구간 유의성을 검정하였다.
결과 및 고찰
생육특성
일미찰옥수수의 생장시기(초엽, 제 1엽, 제 2엽)별, 저온처리 기간(3, 5, 7일)과 온도(5, 3, 0, -3°C)에 따른 저온처리 및 처리 후의 초장의 변화를 나타낸 결과는 Table 1과 같다. 초엽기에서는 저온처리 3일차에 대조구를 제외한 모든 온도 처리구에서 유의성이 없었으나, 다른 처리구에서 저온기간이 길어질수록 온도별 유의성이 뚜렷하게 나타났다. 5°C 이하 5일 이상 처리시 온도가 낮아질수록 제 1엽기는 63%, 제 2엽기에서는 32% 이상의 생장 저해가 뚜렷하게 나타났다. 처리 후 회복기에서는 초장의 변화가 나타났으나, 초엽기 및 제 1엽기에서 3°C 이상 7일 처리구, 제 2엽기 0°C 이하 7일 처리구는 초장의 차이가 없었다. 특히, 제 2엽기의 0°C 이하 7일 처리구와 모든 생장시기의 –3°C 처리구에서는 처리 후, 회복기 전·후 초장의 변화가 없는 것으로 조사되었다.
Table 1.
Changes in plant height according to the chilling treatment period and temperature by growth stage.
| Chilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | cm | % | cm | % | cm | % | cm | % | cm | % | cm | % | |||||
| Before | 6.4 | 9.2 | 15.0 | |||||||||||||||
| 3 | Con. | 23.7a‡ | 0 | 26.7a | 0 | 26.7a | 0 | 43.3a | 0 | 51.8a | 0 | 58.6a | 0 | |||||
| 5 | 6.9b | 71 | 12.2b | 64 | 20.9b | 22 | 30.7b | 29 | 32.7b | 37 | 56.7b | 3 | ||||||
| 3 | 7.1b | 70 | 11.8b | 56 | 17.5c | 34 | 28.0c | 35 | 29.0c | 44 | 41.5c | 29 | ||||||
| 0 | 7.2b | 70 | 10.2c | 62 | 15.3c | 43 | 25.7d | 41 | 23.7d | 54 | 25.6d | 46 | ||||||
| -3 | 6.4b | 73 | 9.2c | 66 | 15.0c | 44 | 6.4e | 85 | 9.2e | 82 | 15.0e | 74 | ||||||
| 5 | Con. | 31.1a | 0 | 38.1a | 0 | 39.0a | 0 | 44.9a | 0 | 56.8a | 0 | 70.4a | 0 | |||||
| 5 | 10.3b | 67 | 14.2b | 63 | 26.5b | 32 | 20.8b | 54 | 32.3b | 43 | 53.2b | 24 | ||||||
| 3 | 7.3c | 77 | 12.8c | 66 | 23.5c | 40 | 19.0c | 58 | 27.8c | 51 | 34.5c | 51 | ||||||
| 0 | 7.5c | 76 | 10.5d | 72 | 16.2d | 58 | 18.1d | 60 | 23.7d | 58 | 23.5d | 67 | ||||||
| -3 | 6.4d | 79 | 9.2e | 76 | 15.0e | 62 | 6.4e | 86 | 9.2e | 84 | 15.0e | 79 | ||||||
| 7 | Con. | 42.2a | 0 | 46.4a | 0 | 47.4a | 0 | 49.9a | 0 | 63.1a | 0 | 73.1a | 0 | |||||
| 5 | 12.7b | 70 | 14.7b | 68 | 27.3b | 42 | 16.3b | 67 | 32.3b | 49 | 45.2b | 38 | ||||||
| 3 | 10.1c | 76 | 13.2c | 72 | 25.5c | 46 | 16.2b | 68 | 26.3b | 58 | 25.2c | 61 | ||||||
| 0 | 8.2d | 81 | 11.7d | 73 | 16.3d | 66 | 12.2c | 76 | 18.2c | 71 | 16.0d | 68 | ||||||
| -3 | 6.4e | 85 | 9.2e | 80 | 15.0e | 68 | 6.4d | 87 | 9.2d | 85 | 15.0d | 69 | ||||||
벼의 냉해양상은 못자리에 8~10°C 이하에서 냉해를 받고, 유묘기에 13°C 이하에서 발아 및 생육이 늦어지며, 생장기에 12~13°C 이하에서는 냉해에 의해 초장과 분얼이 감소한다고 하였고(Chae et al., 2009), 벼 신품종별 저온장해연구에서는 주야 10/5°C 처리하였을 때, 제 2엽기에서 4일간 처리시 초장의 생장이 거의 중지되었고, 3일째부터 위조고사가 시작되었다고 보고되었다(Lee et al., 1977). 일미찰옥수수의 경우도 모든 온도 처리구에서 5°C 3일차부터 생장이 지연되었으며, 제 2엽기의 0°C 7일 처리구 및 –3°C 이하 3일째부터 생장이 정지된 것으로 조사되었다.
일미찰옥수수의 생장시기별 저온처리 기간과 온도에 따른 저온처리 및 처리후의 경태 변화를 나타낸 결과는 Table 2와 같다. 먼저, –3°C 모든 처리구에서 처리전 대비 0.52~ 1.70 mm 감소한 것으로 나타났다. 생장시기별 처리기간이 길어지고, 온도가 낮아질수록 경태의 성장이 저해되는 것으로 나타나 특히, 5°C 이하 7일차에서 초엽(21% 이상), 제 1엽기(15% 이상) 및 제 2엽기(29% 이상)에서 저해율이 가장 높게 나타났다. 처리 후 회복기 초엽기에서는 모든 저온기간별 온도 처리구에서 11~65%로 생장 저해율이 가장 낮았다. 반면, 제 2엽기 5°C 이하 7일차에 43~77%로 가장 높은 저해율이 나타났다. 전 생장시기 및 저온기간에서는 온도가 낮아짐에 따라 경태가 더 짧게 나타났고, 저해율이 증가하여 5°C 이하 7일 처리구에서 가장 높은 것으로 조사되었다. 특히 저온 7일차에서 제 2엽기 5°C 처리구를 제외한 모든 생장 시기 및 온도 처리구에서 저온처리 대비 처리 후 회복기에 경태가 오히려 감소하였고, 제 1엽기에서는 0°C 5일차부터 감소하였으며, 제 2엽기에서도 0°C에서는 3일차부터, 3°C에서는 5일차부터 경태가 감소하였다. 또한, –3°C 모든 처리구에서는 경태의 변화가 조사되지 않았다.
Table 2.
Changes in stem dimeter according to the chilling treatment period and temperature by growth stage.
| Cilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | mm | % | mm | % | mm | % | mm | % | mm | % | mm | % | |||||
| Before | 1.96 | 2.56 | 2.84 | |||||||||||||||
| 3 | Con. | 2.85a‡ | 0 | 3.08a | 0 | 3.39a | 0 | 4.00a | 0 | 4.69a | 0 | 4.82a | 0 | |||||
| 5 | 2.37b | 17 | 2.80b | 9 | 2.91b | 14 | 3.54b | 11 | 4.39b | 6 | 4.61a | 4 | ||||||
| 3 | 2.42b | 15 | 2.74bc | 11 | 2.87bc | 15 | 3.18c | 20 | 3.22c | 31 | 3.04b | 37 | ||||||
| 0 | 2.06c | 28 | 2.60c | 16 | 2.83c | 17 | 3.04d | 24 | 2.87d | 39 | 2.50c | 48 | ||||||
| -3 | 1.31d | 48 | 1.44d | 52 | 1.47d | 66 | 1.31e | 63 | 1.44e | 69 | 1.47d | 70 | ||||||
| 5 | Con. | 3.21a | 0 | 3.61a | 0 | 4.06a | 0 | 4.13a | 0 | 5.12a | 0 | 5.59a | 0 | |||||
| 5 | 2.60b | 19 | 3.37b | 7 | 2.97b | 27 | 3.23b | 22 | 3.43b | 33 | 4.23b | 24 | ||||||
| 3 | 2.57b | 20 | 3.04c | 16 | 2.94b | 28 | 3.00c | 27 | 3.05c | 40 | 2.74c | 51 | ||||||
| 0 | 2.14c | 33 | 2.68d | 26 | 2.87c | 29 | 2.99c | 28 | 2.39d | 53 | 2.16d | 61 | ||||||
| -3 | 1.44d | 54 | 1.47e | 59 | 1.47d | 64 | 1.44d | 64 | 1.47e | 71 | 1.47e | 74 | ||||||
| 7 | Con. | 4.00a | 0 | 4.16a | 0 | 4.52a | 0 | 4.23a | 0 | 5.40a | 0 | 6.47a | 0 | |||||
| 5 | 3.14b | 21 | 3.55b | 15 | 3.20b | 29 | 3.13b | 26 | 3.16b | 41 | 3.67b | 43 | ||||||
| 3 | 2.96c | 26 | 3.09c | 26 | 3.18b | 30 | 2.66c | 37 | 2.95c | 45 | 2.18c | 66 | ||||||
| 0 | 2.14d | 47 | 2.84d | 32 | 3.08c | 32 | 2.66c | 37 | 2.06d | 62 | 1.92d | 70 | ||||||
| -3 | 1.16e | 63 | 1.15e | 65 | 1.14d | 67 | 1.16d | 65 | 1.15e | 73 | 1.14e | 77 | ||||||
세포내 결빙은 원형질 구성에 필요한 수분이 동결해 원형질단백이 응고하고 변화해 원형질의 구조가 파괴되므로 세포가 동사하게 되는데(Chae et al., 2009), 위의 -3°C 처리구도 급격한 온도 저하로 인해 세포내 결빙에 의해 고사한 것으로 사료되며, 동결되었을 때 세포안에서 세포간극으로 투과·이동한 수분이 융해시 세포안으로 다시 투과되기 전 증발하여 잎의 수분부족 현상이 나타나게 되며(Chae et al., 2009), 본 연구에서도 이와 같은 원인에 의해 제 1엽기 저온 0°C 5일부터, 제 2엽기에서도 0°C에서는 3일, 3°C에서는 5일부터 경태가 감소하였으며, –3°C 이하 3일부터 생장이 정지된 것으로 조사되었다.
Table 3에서도 –3°C 처리구에서는 저온처리 및 처리 후 회복기의 모든 처리구에서 엽수의 변화가 없는 것으로 조사되었고, 제 2엽기에서도 저온처리별 엽수의 변화가 나타나지 않았다. 초엽기와 제 1엽기에서는 0°C 이하 3일차 처리구부터 엽수의 변화가 없었다. 처리 후 회복기의 엽수의 변화에서는 먼저, 초엽기와 제 1엽기에서의 3°C 이상 3일 처리구에서는 대조구와의 유의성이 나타나지 않았다. –3°C 모든 처리구와 제 2엽기에서는 3°C 이하 7일 처리구부터 처리 후 회복기에서의 엽수의 변화가 나타나지 않았다.
Table 3.
Changes in leaf numbers according to the chilling treatment period and temperature by growth stage.
| Cilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | ea | % | ea | % | ea | % | ea | % | ea | % | ea | % | |||||
| Before | 0.00 | 1.00 | 2.00 | |||||||||||||||
| 3 | Con. | 1.00a‡ | 0 | 2.00a | 0 | 2.25a | 0 | 3.00a | 0 | 4.00a | 0 | 4.33a | 0 | |||||
| 5 | 0.00b | 100 | 1.67a | 0 | 2.00a | 11 | 3.00a | 0 | 3.67a | 8 | 4.00ab | 8 | ||||||
| 3 | 0.00b | 100 | 1.50b | 25 | 2.00a | 11 | 2.67a | 11 | 3.33a | 17 | 4.00ab | 8 | ||||||
| 0 | 0.00b | 100 | 1.00c | 50 | 2.00a | 11 | 2.00b | 33 | 2.33b | 42 | 3.33b | 23 | ||||||
| -3 | 0.00b | 100 | 1.00c | 50 | 2.00a | 11 | 0.00c | 100 | 1.00c | 75 | 2.00c | 54 | ||||||
| 5 | Con. | 2.00a | 0 | 3.00a | 0 | 3.67a | 0 | 3.67a | 0 | 4.00a | 0 | 4.67a | 0 | |||||
| 5 | 1.00b | 50 | 1.67b | 44 | 2.00b | 46 | 2.67b | 27 | 3.33b | 17 | 4.00a | 14 | ||||||
| 3 | 1.00b | 50 | 1.50b | 56 | 2.00b | 46 | 2.00c | 46 | 3.00b | 25 | 4.00a | 14 | ||||||
| 0 | 0.00c | 100 | 1.00b | 67 | 2.00b | 46 | 1.00d | 73 | 2.00c | 50 | 2.67b | 43 | ||||||
| -3 | 0.00c | 100 | 1.00b | 67 | 2.00b | 46 | 0.00e | 100 | 1.00d | 75 | 2.00c | 57 | ||||||
| 7 | Con. | 2.67a | 0 | 4.00a | 0 | 4.33a | 0 | 4.00a | 0 | 4.67a | 0 | 4.67a | 0 | |||||
| 5 | 1.00b | 63 | 1.67b | 75 | 2.00b | 54 | 2.33b | 42 | 2.67b | 43 | 3.67b | 21 | ||||||
| 3 | 1.00b | 63 | 1.50b | 75 | 2.00b | 54 | 2.00b | 50 | 2.33bc | 50 | 2.67c | 43 | ||||||
| 0 | 0.00c | 100 | 1.00b | 75 | 2.00b | 54 | 0.67c | 83 | 1.67cd | 64 | 2.00c | 57 | ||||||
| -3 | 0.00c | 100 | 1.00b | 75 | 2.00b | 54 | 0.00d | 100 | 1.00d | 79 | 2.00c | 57 | ||||||
생장시기별 저온처리 기간과 온도에 따른 저온처리 및 처리 후의 엽색도(SPAD) 변화에서는 –3°C 처리구의 모든 온도 처리구에서 9.2~19.8로 낮게 조사되었고, 생장저해율도 50% 이상으로 조사되었으며, 특히, 저온처리 및 처리 후 회복기의 엽색도의 변화가 없었다(Table 4). 모든 생장시기에서 저온기간이 증가하거나 온도가 낮아질수록 엽색도 값이 낮아지는 경향을 보였고, 초엽기 및 제 2엽기에서는 저온 5일 이상에서 온도에 대한 차이가 뚜렷하게 조사되었다. 저해율에서는 초엽기의 0°C 3일 처리구, 3°C 7일 처리구부터 30% 이상으로 조사되었고, 제 2엽기에서는 0°C 3일 처리구와 3°C 5일 처리구부터 30% 이상으로 조사되어 생장시기가 늦어질수록, 저온기간이 길어질수록, 온도는 낮아질수록 저해율은 높아지는 것으로 나타났다. 처리 후 회복기에서는 초엽기 5°C 7일차 처리구에서도 SPAD 30 이상으로 나타났으나, 제 2엽기 –3°C 5일차부터는 SPAD 10 이하로 나타났다. 대조구 대비 성장 저해율에서는 제 2엽기 –3°C 7일 처리구가 80%로 가장 높았으나, 초엽기 5°C 3일 처리구에서 14%로 가장 낮게 나타났다. 이처럼 처리 후에서도 생장시기가 늦어질수록, 저온기간이 길어질수록, 온도는 낮아질수록 저해율이 높아지는 것으로 나타났다.
Table 4.
Changes in SPAD according to the chilling treatment period and Temperature by growth stage.
| Cilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | cm | % | cm | % | cm | % | cm | % | cm | % | cm | % | |||||
| Before | 38.6 | 34.8 | 39.2 | |||||||||||||||
| 3 | Con. | 38.9a‡ | 0 | 40.0a | 0 | 40.0a | 0 | 40.6a | 0 | 40.4a | 0 | 42.4a | 0 | |||||
| 5 | 37.8ab | 3 | 34.8a | 13 | 34.0b | 15 | 35.0b | 14 | 29.6b | 27 | 28.8b | 32 | ||||||
| 3 | 37.0b | 5 | 34.5a | 14 | 28.7c | 28 | 27.7c | 32 | 26.2c | 35 | 23.0c | 46 | ||||||
| 0 | 26.3c | 32 | 30.8b | 23 | 25.4c | 37 | 26.2c | 35 | 25.3c | 37 | 21.4d | 50 | ||||||
| -3 | 18.9d | 51 | 17.9c | 55 | 19.8d | 51 | 18.9d | 53 | 17.9d | 56 | 19.7e | 54 | ||||||
| 5 | Con. | 39.4aa | 0 | 40.4a | 0 | 41.0a | 0 | 41.5a | 0 | 41.1a | 0 | 43.6a | 0 | |||||
| 5 | 34.3b | 13 | 28.6b | 29 | 34.5b | 16 | 32.6b | 21 | 26.9b | 35 | 25.7b | 41 | ||||||
| 3 | 31.8c | 19 | 27.5c | 32 | 25.4c | 38 | 27.3c | 34 | 25.6c | 38 | 23.6c | 46 | ||||||
| 0 | 24.2d | 39 | 27.4c | 32 | 24.5d | 40 | 24.8d | 40 | 25.2c | 39 | 22.0d | 50 | ||||||
| -3 | 18.7e | 53 | 17.5d | 57 | 9.7e | 76 | 18.7e | 55 | 17.5d | 57 | 9.4e | 78 | ||||||
| 7 | Con. | 40.3aa | 0 | 40.7a | 0 | 41.3a | 0 | 42.1a | 0 | 42.4a | 0 | 46.4aa | 0 | |||||
| 5 | 32.2b | 20 | 28.2b | 31 | 31.7b | 23 | 30.3b | 28 | 25.8b | 39 | 23.9b | 48 | ||||||
| 3 | 28.3c | 30 | 27.0b | 34 | 25.7c | 38 | 25.0c | 41 | 24.9c | 41 | 20.6c | 56 | ||||||
| 0 | 22.1d | 45 | 27.0b | 34 | 22.2d | 46 | 24.4c | 42 | 24.7c | 42 | 18.2d | 61 | ||||||
| -3 | 18.6e | 54 | 16.5c | 59 | 9.2e | 78 | 18.6d | 56 | 16.5d | 61 | 9.2e | 80 | ||||||
따라서 생장시기가 늦어질수록, 저온기간이 길어질수록, 온도가 낮아질수록 더 많은 스트레스를 받는 것으로 생각되며, 특히, –3°C 모든 생장시기에서와 제 2엽기 0°C 7일차에서는 SPAD 20 이하로 저온에 따른 많은 스트레스를 받은 것으로 생각된다.
식생지수로서 녹색식물의 지표로 이용되고 있는 NDVI 변화(Shin & Kim, 2003)에서도 위의 엽색도 변화와 유사한 경향으로 모든 생장시기에서 저온기간이 길어지고, 온도가 낮아질수록 값이 낮아지는 경향으로 나타났다(Table 5). 대조구 대비 처리에 따른 저해율은 제 2엽기에서 높게 나타났으며, 제 2엽기>제 1엽기>초엽기 순으로 높은 저해율이 나타났다. 특히, 제 2엽기 5°C 이하 7일 처리구에서 0.142~0.686으로 가장 낮게 조사되었으며, 이 중 0°C 처리구에서 0.613으로 대조구 대비 21% 의 저해율로 –3°C 처리구 제외한 처리구 중 가장 낮은 것으로 조사되었다. 처리 후 회복기의 저해율에서는 제 2엽기 5°C 이하 7일 처리구에서 25~82%로 가장 높았으며, 초엽기 5°C 3일 처리구에서 4%로 가장 낮았다. 모든 처리구의 –3°C 처리에서는 대조구 대비 61% 이상으로 저해율이 높게 나타났고, 저온처리 대비 NDVI 값이 증가하지 않았다.
Table 5.
Changes in NDVI according to the chilling treatment period and temperature by growth stage.
| Cilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | cm | % | cm | % | cm | % | cm | % | cm | % | cm | % | |||||
| Before | 0.740 | 0.760 | 0.770 | |||||||||||||||
| 3 | Con. | 0.742a‡ | 0 | 0.754a | 0 | 0.770a | 0 | 0.747a | 0 | 0.771a | 0 | 0.784a | 0 | |||||
| 5 | 0.733a | 1 | 0.707b | 6 | 0.731b | 5 | 0.715b | 4 | 0.730b | 5 | 0.672b | 14 | ||||||
| 3 | 0.716b | 4 | 0.681c | 10 | 0.716b | 7 | 0.700b | 6 | 0.643c | 17 | 0.627c | 20 | ||||||
| 0 | 0.634c | 15 | 0.671c | 11 | 0.696c | 10 | 0.677c | 9 | 0.605d | 22 | 0.569d | 27 | ||||||
| -3 | 0.290d | 61 | 0.182d | 76 | 0.150d | 81 | 0.222d | 70 | 0.182e | 76 | 0.150e | 81 | ||||||
| 5 | Con. | 0.736a | 0 | 0.764a | 0 | 0.770a | 0 | 0.750a | 0 | 0.780a | 0 | 0.800a | 0 | |||||
| 5 | 0.710a | 4 | 0.737b | 4 | 0.692b | 10 | 0.703b | 6 | 0.685b | 12 | 0.617b | 23 | ||||||
| 3 | 0.639b | 13 | 0.686c | 10 | 0.692b | 10 | 0.648c | 14 | 0.530c | 32 | 0.584c | 27 | ||||||
| 0 | 0.621b | 16 | 0.670c | 12 | 0.661c | 14 | 0.597d | 20 | 0.504d | 35 | 0.559c | 30 | ||||||
| -3 | 0.222c | 70 | 0.158d | 79 | 0.137d | 82 | 0.222e | 70 | 0.158e | 80 | 0.137d | 83 | ||||||
| 7 | Con. | 0.735a | 0 | 0.769a | 0 | 0.772a | 0 | 0.740a | 0 | 0.781a | 0 | 0.803a | 0 | |||||
| 5 | 0.635b | 14 | 0.743a | 3 | 0.686b | 11 | 0.704b | 5 | 0.625b | 20 | 0.604b | 25 | ||||||
| 3 | 0.631b | 14 | 0.674b | 12 | 0.637c | 17 | 0.569c | 23 | 0.505c | 35 | 0.521c | 35 | ||||||
| 0 | 0.620c | 16 | 0.670b | 13 | 0.613d | 21 | 0.558c | 25 | 0.490d | 37 | 0.504d | 37 | ||||||
| -3 | 0.216d | 71 | 0.151c | 80 | 0.142e | 82 | 0.216d | 71 | 0.151e | 81 | 0.142e | 82 | ||||||
따라서 이러한 결과는 Lee et al. (1977)에 따르면 벼의 유묘기에 2엽기 주야 10/5°C 저온처리시 일부 품종에서 저온처리 직후에 10% 내외 고사율을 보였지만, 처리완료 후 고온에 환원하여도 회복되지 못하고 고사율이 증가했다는 보고와 같이 NDVI 값도 저온처리 보다 처리 후 측정한 값에서 더 낮게 조사되어 유사한 결과로 나타났다. 또한, 대조구 대비 처리에 따른 생장 저해율이 제 2엽기>제 1엽기>초엽기 순으로 나타나, 생장 시기가 길어질수록 높은 스트레스를 받는 것으로 생각되며, 특히 -3°C 처리구에서 처리 전 및 대조구 대비 61~82%로 높은 저해율이 나타났고, 처리 후 회복기에서도 변화가 없는 것으로 조사되어, Koo et al.(2003)이 보고한 –2~–3°C의 저온조건을 저온항온기에서 5~6시간만 유지시켜도 식물체 세포내 결빙을 유도한다고 한 결과에서와 같이 본 연구에서도 세포내 결빙에 의해 유묘가 고사한 것으로 사료된다.
Table 6과 같이 식물의 광합성 수행 능력의 지표로 이용되고 있는 변동형광값/최대형광값비(Fv/Fm)의 변화에서는(Lee et al., 2014) –3°C 처리구가 모든 생장시기별 온도 처리구에서 0.099 이하로 조사되었고, 처리 후 회복기에서도 값의 변화는 나타나지 않았다. 전 생장시기별 5°C 3일 처리구에서 대조구와 차이가 없었으나, 이후 모든 처리구에서 대조구 대비 변화가 조사되었다. 대조구 대비 처리에 따른 저해율은 제 2엽기에서 3~98%로 가장 높게 나타났으며, 앞의 NDVI 변화에서와 같이 제 2엽기>제 1엽기>초엽기 순으로 높은 저해율이 나타났다. 처리 후 회복기에서 모든 생장시기별 5°C 이하 7일 처리구에서 7~98%로 가장 높은 저해율을 보였고, 초엽기 및 제 1엽기 5°C 3일 처리구에서 저온처리와 동일하게 대조구와 차이가 없었던 반면, 제 2엽기 5, 3°C 이상 3일 처리구에서는 0.747, 0.704로 저온처리 대비 각각 0.058, 0.083 감소하여 대조구 대비 10~15%의 저해율이 나타나, 생장시기와 처리 기간이 길어질수록 온도가 낮아질수록 값이 낮아지고, 저해율은 증가하는 것으로 조사되었다.
Table 6.
Changes in Fv/Fm according to the chilling treatment period and temperature by growth stage.
| Cilling treatment | After treatment | |||||||||||||||||
| Coleoptile | 1st leaf | 2nd leaf | Coleoptile | 1st leaf | 2nd leaf | |||||||||||||
| I.R. | I.R. | I.R. | I.R. | I.R. | I.R. | |||||||||||||
| days | °C | % | % | % | % | % | % | |||||||||||
| Before | 0.820 | 0.820 | 0.830 | |||||||||||||||
| 3 | Con. | 0.824a‡ | 0 | 0.817a | 0 | 0.831a | 0 | 0.840a | 0 | 0.830a | 0 | 0.832a | 0 | |||||
| 5 | 0.812a | 1 | 0.806a | 1 | 0.805a | 3 | 0.823a | 2 | 0.824a | 1 | 0.747b | 10 | ||||||
| 3 | 0.776b | 6 | 0.783ab | 4 | 0.787a | 5 | 0.747b | 11 | 0.747b | 10 | 0.704c | 15 | ||||||
| 0 | 0.739c | 10 | 0.764b | 6 | 0.693b | 17 | 0.726b | 14 | 0.677c | 18 | 0.616d | 26 | ||||||
| -3 | 0.076d | 91 | 0.099c | 88 | 0.086c | 90 | 0.076c | 91 | 0.099d | 88 | 0.086e | 90 | ||||||
| 5 | Con. | 0.829a | 0 | 0.822a | 0 | 0.832a | 0 | 0.843a | 0 | 0.831a | 0 | 0.837a | 0 | |||||
| 5 | 0.784b | 5 | 0.796b | 3 | 0.778b | 6 | 0.808b | 4 | 0.797b | 4 | 0.713b | 15 | ||||||
| 3 | 0.760c | 8 | 0.681c | 17 | 0.643c | 23 | 0.729c | 14 | 0.717c | 14 | 0.653c | 22 | ||||||
| 0 | 0.734d | 11 | 0.677c | 18 | 0.641c | 23 | 0.622d | 26 | 0.623d | 25 | 0.606d | 28 | ||||||
| -3 | 0.046e | 94 | 0.033d | 96 | 0.063d | 92 | 0.046e | 95 | 0.033e | 96 | 0.063e | 92 | ||||||
| 7 | Con. | 0.836a | 0 | 0.828a | 0 | 0.832a | 0 | 0.843a | 0 | 0.827a | 0 | 0.840a | 0 | |||||
| 5 | 0.775b | 7 | 0.766b | 7 | 0.792b | 5 | 0.787b | 7 | 0.721b | 13 | 0.709b | 16 | ||||||
| 3 | 0.689c | 18 | 0.657c | 21 | 0.642c | 23 | 0.711c | 16 | 0.630c | 24 | 0.617c | 27 | ||||||
| 0 | 0.640d | 23 | 0.630d | 24 | 0.609d | 27 | 0.616d | 27 | 0.605d | 27 | 0.530d | 37 | ||||||
| -3 | 0.037e | 96 | 0.020e | 98 | 0.020e | 98 | 0.037e | 96 | 0.020e | 98 | 0.020e | 98 | ||||||
따라서 이러한 결과는 Lee et al. (1977)의 연구결과와 유사한 경향으로 회복 중 저온스트레스를 받은 것으로 생각된다. 앞의 NDVI 변화에서도 유사한 결과로 생장시기와 처리기간이 길어질수록, 온도가 낮을수록 저해율은 증가하여 저온 스트레스로 인한 광합성 수행능력이 더 낮아진 것으로 사료되며, 특히 –3°C 처리구에서는 Fv/Fm 값에서 0.09 이하로 나타나고, 처리 후 회복에서도 변화가 없는 것으로 조사되어, 생장이 정지된 것으로 사료된다. 또한, 초엽기의 5°C 3일 처리구에서 NDVI와 Fv/Fm 값이 대조구와 차이가 없어, 서로 같은 경향을 보였다. SPAD 및 NDVI 값에서는 저온처리 대비 처리 후에 값이 감소하는 경향을 보인 반면, Fv/Fm 값에서는 일부 처리에서 증가하는 경향을 보여 향후, 저온처리에 따른 SPAD 및 NDVI 값과 Fv/Fm 값의 상관관계를 조사하는 연구가 추가로 이루어지면 좋을 것으로 사료된다.
저온 생장저해 양상
생장시기에 따른 저온기간별 온도 처리구에서 피해양상을 조사한 결과는 Fig. 1과 같다. 피해양상은 위조된 엽의 면적 및 개수를 지수로 환산한 결과로 –3°C 처리구에서는 모든 처리구에서 지수 9로 가장 높게 나타났다. 초엽기에서는 저온기간별 3°C 이상 처리구에서 지수 0으로 나타났고, 처리기간이 길어질수록 변화는 나타나지 않았다. 제 1엽에서는 모든 온도 처리구에서 7일차에 지수 2 이상으로 조사되었으며, 제 2엽기에서도 모든 온도 처리구에서 7일차에 지수 2 이상의 피해양상이 나타났다. 제 2엽기 저온 0°C 7일 처리구에서는 초엽기(지수 2) 대비 3배 많은 지수 6으로 조사되었다.
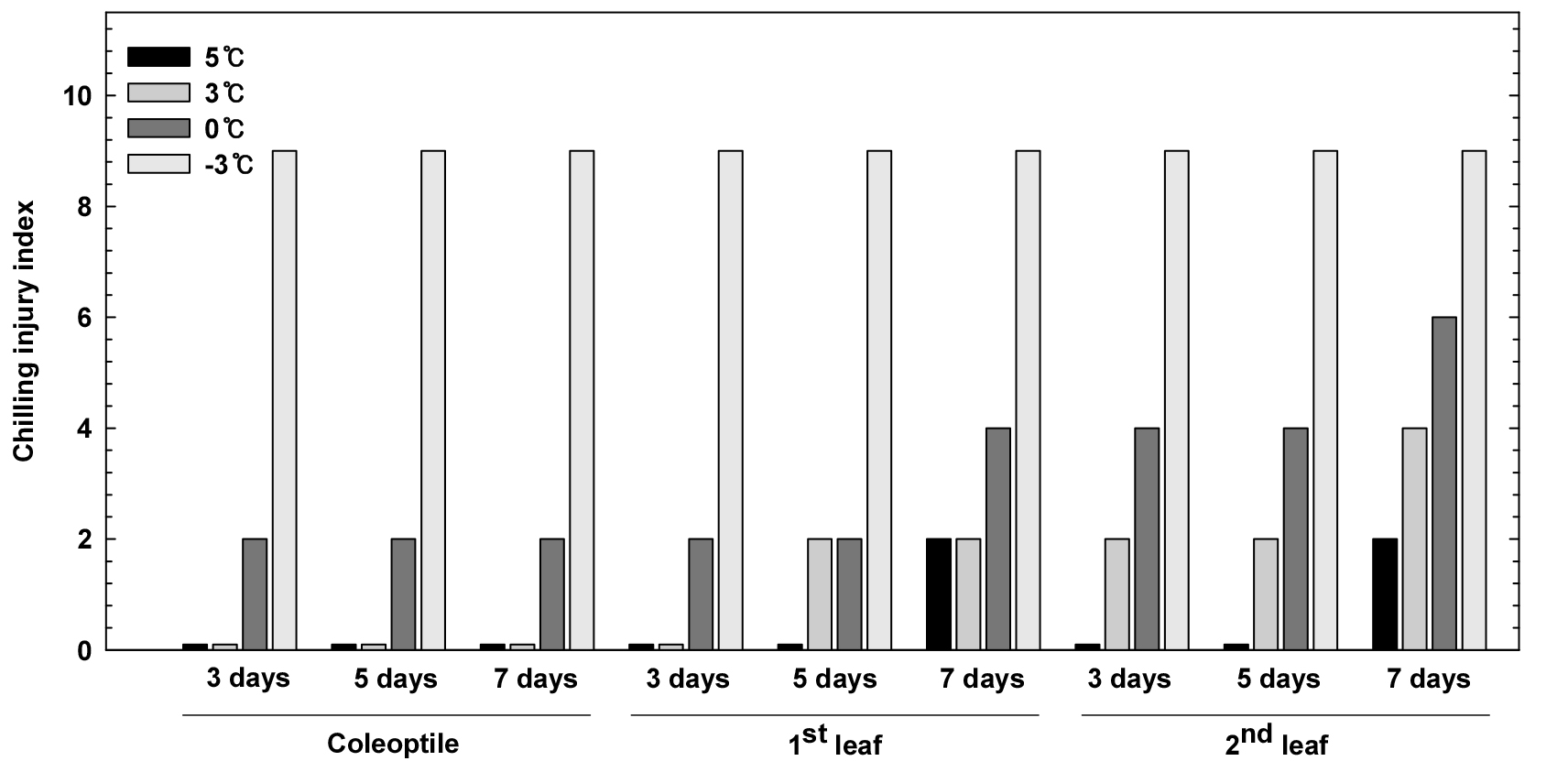
따라서 이러한 결과는 Koo et al. (2003)이 보고한 맥류 유수발육기 저온실험시 –3°C 정도의 저온조건이 퇴화, 불임 등 장해증상을 보다 정밀하게 조사가능하다고 하였으며, 저온유지시간이 장해발생에 중요요인이라고 언급한 것처럼 본 연구에서도 –3°C에서 생육정지 및 위조고사가 나타나 피해양상이 가장 높게 나타났고, 저온기간이 길어질수록 같은 온도 처리구라도 지수가 더 높게 나타나 피해양상이 증가하였다. 따라서 초엽 및 제 1엽 대비 제 2엽에서 저온에 대한 피해양상이 더 도드라졌으며, 저온기간이 길어질수록 피해가 증가한 것으로 사료된다.
상관관계
생장시기별 온도 및 저온기간에 따른 생육특성과의 상관관계를 분석한 결과 생장시기에서는 초장 및 엽수에서 각각 0.310**, 0.475**으로 높은 양의 상관관계가 나타났다(Table 7). 온도와 생육특성과의 상관관계에서는 초장(-0.647**), 경태(-0.796**), 엽수(-0.543**), SPAD (-0.093**), NDVI (-0.844**) 및 Fv/Fm (-0.836**)에서 모두 높은 음의 상관관계가 나타났다. 저온기간과 생육특성과의 상관관계에서는 SPAD에서 –0.178**으로 높은 음의 상관관계가 나타났고, NDVI 및 Fv/Fm에서도 각각 –0.141*, -0.132*으로 상관관계가 나타났다.
Table 7.
Correlation coefficients between the growth stage, chilling period, and temperature.
| Growth period | Temperature | Chilling period | |
| Plant height | 0.310** | -0.647** | 0.085ns |
| Stem D | 0.134* | -0.796** | 0.091ns |
| Leaf n | 0.475** | -0.543** | 0.054ns |
| SPAD | -0.146* | -0.093** | -0.178** |
| NDVI | -0.042ns | -0.844** | -0.141* |
| Fv/Fm | -0.046ns | -0.836** | -0.132* |
따라서 위의 내용을 종합한 결과 모든 생장시기에서 초장, 경태 및 엽수뿐만 아니라 식생지수(NDVI) 및 광합성 능력지수(Fv/Fm)에서도 저온 기간이 길어지고, 온도가 낮아짐에 따라 대조구 대비 저온 스트레스가 증가하여, 저해율도 높은 값이 나타났다. 특히, 식생지수 및 광합성 능력지수에서는 제 2엽기에서 높은 저해율이 나타났으며, 제 2엽기>제 1엽기>초엽기 순으로 높은 저해율을 나타나, 뚜렷한 경향을 보였다. 또한, 처리후 회복기에서는 모든 처리구의 –3°C 처리구 및 제 2엽기에서는 0°C 7일 처리구부터 변화가 없는 것으로 조사되어, Koo et al. (2003)이 보고한 –2~–3°C의 저온조건을 저온항온기에서 5~6시간만 유지시켜도 식물체 세포내 결빙을 유도한다는 결과와 같이 본 연구에서도 일미찰옥수수의 전 유묘기에는 –3°C에서는 3일부터, 0°C에서는 제 2엽기 7일부터 세포내 결빙에 의해 유묘가 고사한 것으로 사료된다.
적 요
본 연구는 남부지방에서 찰옥수수와 콩의 2모작 직파 표준재배법 확립을 위한 기초자료로 활용하고자 찰옥수수의 유묘기 저온에 따른 생육특성 변화 및 피해양상을 조사하였으며 그 결과는 다음과 같다
일미찰옥수수 초장은 5°C 이하 3일 처리구부터 저해가 나타나 모든 처리구에서 22% 이상의 저해율이 조사되었다. 경태도 저온기간이 길어지고, 온도가 낮아질수록 더 짧은 것으로 나타나, 제 2엽기 5°C 이하 5일 처리구에서 저해율이 27% 이상으로 가장 높게 나타났다. 식생지수(NDVI)에서 저해율은 제 2엽기>제 1엽기>초엽기 순으로 높게 나타났으며, 회복기 저해율에서는 제 2엽기 5°C 이하 7일 처리구에서 25~82%로 가장 높았다. 광합성능력지수(Fv/Fm)에서도 제 2엽기>제 1엽기>초엽기 순으로 저해율이 높게 조사되었다. 특히, –3°C 처리구의 저온처리 및 회복기에서 생육특성의 변화가 없거나, 감소하는 것으로 조사되었다. 따라서 5°C 이하 3일부터 저온기간이 길어지고, 온도가 낮아질수록 생장이 저해되었고, 그 피해정도는 제 2엽기>제 1엽기>초엽기 순으로 높았다. 모든 유묘기에서 –3°C 이하에 3일 이상 노출될 경우 생육이 정지되거나, 위조고사 하였다. 한편, 본 실험은 실내에서 수행된 결과이므로 차후 재배지역 적용여부에 대한 면밀한 추가 연구가 수행되어야 할 것으로 사료된다.
