

서 론
재료 및 방법
온도구배챔버(Temperature Gradient Chambers: TGCs)
재배 및 수량구성요소 조사
종실 단백질과 지방함량 분석
결 과
온도와 생물 계절 변화
수량구성요소와 수량 변화
온도 상승과 종실 단백질 및 지방함량 변화
고 찰
생물계절 및 착협수 변화
종실 크기와 수량 변화
종실 단백질과 지방함량 변화
적 요
서 론
2018년 기후변화에 관한 정부간 패널(Intergovernmental Panel on Climate Change: IPCC, 2018)에 의하면 현재 속도로 지구온난화가 지속된다면 2030-2052년 사이 약 1.5°C가 상승할 것으로 보고되었다.
온도는 작물의 생장과 발달에 매우 주요한 요인 중에 하나이다(Yamori et al., 2005). 그러나 작물재배기간 고온은 작물 생산성에 부정적인 영향을 준다(Dajanaguiraman et al., 2013; Wheeler & von Braun, 2013). 생육기간 고온은 생육 발달과 개화기 지연 그리고 생화학적 반응으로 단백질 변성 유발과 세포막 지질 유동성 및 활성산소가 증가를 하고, 이로 인해 광합성과 관련된 조직의 기능을 약화시킨다 하였다(Larkindale et al., 2005; Hasanuzzaman et al., 2013).
콩은 대표적인 기름 작물로서 식용, 사료용 및 산업용에 걸쳐 우리 주변에서 다양한 용도로 이용되고 있다. 2018년 전국 콩 재배면적은 5만 638 ha로 2017년 4만 5.556 ha 보다 약 11%가 증가하였다(Ha, 2019). 같은 기간 생산량은 8만 9.410 ton으로 2017년 8만 5.644 ton보다 약 4.4%가 증가하였다. 그러나 재배 면적과 생산량 증가에도 불구하고, 단위면적당 수량은 크게 증가하지 않고 있다. 이러한 원인은 다양하지만, 최근 지구온난화로 인한 폭우와 폭염과 같은 비생물적 스트레스의 발생 빈도가 높아지면서 수량 증가에 저해를 초래하고 있다.
우리나라 콩은 일반적으로 늦봄에 파종하여 가을에 수확을 한다. 개화기는 7월 하순에서 8월 하순 사이로 이 기간은 생육기간 중 온도가 가장 높은 시기이다. 콩의 수량은 연차간 기상 환경 변화에 매우 민감하게 반응하는데, 특히 생식생장기의 고온은 작물 생육 발달과 수량에 매우 부정적인 영향을 준다 하였다(Ergo et al., 2018).
콩의 생육 적정 온도 범위는 25-30°C (Hesketh et al., 1973; Thuzar et al., 2010)로 평균 온도가 20°C이하 이거나 35°C이상에서는 수량 27%가 감소한다 하였다(Giboson & Mullen, 1996). 콩 개화는 일장에 따라 반응하지만 생육 적정 온도 범위 보다 약간 높으면 개화가 일찍 일어난다 하였다(Garner & Allard, 1930; Egli & Bruening, 1992). 그러나 30°C이상의 고온에 장기간 노출되면 개화기가 지연되면서 종실비대기간도 상당히 지연된다 하였다(Oh-E et al., 2007; Tacarindud et al., 2013). 개화기의 고온은 꽃 가루의 수분 능력이 떨어지고, 꽃 수가 줄고 불규칙한 개화 현상과 함께 폐화 수정이 증가한다 하였다(Dajanaguiraman et al., 2013; Sage et al., 2015; Sita et al., 2017). 결국 고온은 적정 온도에 비하여 착협률과 착협수를 감소한다 하였다(Dajanaguiraman et al., 2013; Allen et al., 2018). 또한 등숙기간 고온은 적정 온도에 비하여 종실 무게를 급격하게 감소시킨다 하였다(Thomas et al., 2010; Tacarindua et al., 2013; Choi et al., 2016).
콩의 종실에는 약 40%의 단백질과 20%의 지방함량을 함유하고 있다(Bellaloui et al., 2011). 대부분의 콩 종실 성분은 환경 변화에 따라 매우 민감하게 반응을 한다(Song et al., 2016). 평균 온도 26-28°C에서는 단백질함량은 증가하지만 지방함량은 이 범위를 벗어나게 되면 감소한다 하였다(Doornbos & Mullen, 1992). 그러나 등숙기간 40°C를 넘게 되면 단백질과 지방함량 모두 감소한다 하였다(Thomas et al., 2003).
따라서 본 시험은 지구온난화에 대한 대응 목적으로 재배기간동안 온도 상승이 생물 계절, 수량구성요소, 단백질 및 지방함량 변화에 대해서 구명하자 포장 환경과 가장 유사한 온도구배 챔버(Temperature Gradient Chambers: TGCs) (Horie et al., 1995)에서 2년간 시험을 수행하였다.
재료 및 방법
온도구배챔버(Temperature Gradient Chambers: TGCs)
본 연구는 국립식량과학원(35° 84' 34" N, 127° 04' 84" E)에 설치된 온도구배챔버에서 수행하였다. TGCs는 가로 2.5 m × 세로 25 m로 외부의 덮개는 폴리올레핀(Polyolefin)으로 피복을 하였다. TGCs는 반 폐쇄형 구조로서 입구는 개방형이고, 안쪽은 환기 팬 2개가 설치되어 있어서 폐쇄되어 있다. 공기와 온도의 흐름은 환기 팬과 열풍기에 의해 일정한 온도구배를 유지하게 설정되어 있다. 따라서 입구부터 외부 온도 대비 약 1°C씩 높아지면서(aT+1°C, aT+2°C, aT+ 3°C, aT +4°C) 가장 안쪽은 외부 온도 대비 약 4°C가 높게 유지된다. 온도는 각 구간 마다 1.5 m 높이에 온도 센서(SP- 110, Apogee Instruments Inc., UT, USA)를 설치하여 기록을 하였다.
재배 및 수량구성요소 조사
시험은 2017년과 2018년에 각각 6월 20일과 6월 22일 TGCs 3개동에서 파종하였다. 시험 품종은 국내에서 가장 많이 재배되고 있는 대원콩, 풍산나물콩 및 대풍콩으로 수행하였다. TGCs간 온도 편차는 ±0.3°C이었다. TGCs의 토성은 모래 54.6%, 실트 30.0% 및 점토 15.4%로 구성된 사양토이다. 재식 거리는 50 cm × 15 cm로 2립씩 파종하였다. 시비량은 N-P2O5-K2O를 10 a당 3-3-3.4 kg으로 파종 전에 전량 기비하였다. 점적호스를 설치하여 파종 후부터 착협기까지 주 2회 그리고 착협기부터 성숙기까지 주 1회 관수 하였다.
생물 계절(Fehr & Caviness, 1977)을 기준으로 개화기(R1), 착협기(R3), 종실비대기(R5) 그리고 성숙기(R7)로 구분하여 조사하였다. 수량 구성요소를 위해 수확기에 처리당 당 20주를 채취해서 주경 마디와 협수 및 100립 중을 조사한 후 종실 수량을 구하였다.
종실 단백질과 지방함량 분석
단백질과 지방함량 분석은 수확된 종실을 75°C의 건조 오븐에서 4일간 건조 처리 후 종실을 곱게 분쇄하여 2 mm 채로 걸렀다. 분쇄된 시료 0.2 g을 Dumass 방법으로 원소 분석기(Elementary, vario MAX cube, GmbH, Germany)를 이용하여 질소 농도를 측정하였다(Fred & Watts, 1993). 단백질 함량은 질소 농도에 단백질 계수 6.25를 곱하여 계산하였다. 지방함량은 분쇄된 시료 3.0 g을 원통여지(Thimble filter)에 넣고 자동 Soxhlet 지방 추출 분석기(Foss, Soxtec 8000, Hilleroed, Denmark)로 180분 정도 추출하였다. 수기에 추출된 지방을 105°C 건조기에 1시간 건조 후 데시케이터에 30분간 방냉 한 후 수기의 지방 무게를 측정하여 지방함량을 구하였다. 각 처리당 단백질과 지방함량은 3반복으로 분석하였다.
통계 분석은 SAS 9.2를 이용하여 ANOVA 분석으로 연차, 품종, 온도구배간 그리고 연차 × 품종 × 온도구배간 상호 분석을 하였다. 그룹간 유의성(P<0.05) 분석은 Tukey 검정으로 하였다.
결 과
온도와 생물 계절 변화
2017년과 2018년 재배기간 일평균 온도, 주간과 야간 온도는 급격하게 차이를 보였다(Fig. 1). 2017년과 2018년의 출현기에서 개화기사이 모든 온도구배의 평균온도는 30°C 이상을 넘었다. 2017년과 2018년 개화기에서 착협시까지 평균온도 범위는 28.2-34.2°C이었다. 그러나 같은 기간 주야간 일일 평균온도는 2018년이 2017년에 비하여 약 3°C이상 높았다. 2017년 주간과 야간 온도는 착협기부터 낮아졌지만, 2018년은 종실비대기 중반까지 주야간 온도가 25°C 이상을 유지하다가 종실비대기 중반 이후부터 낮아졌다.
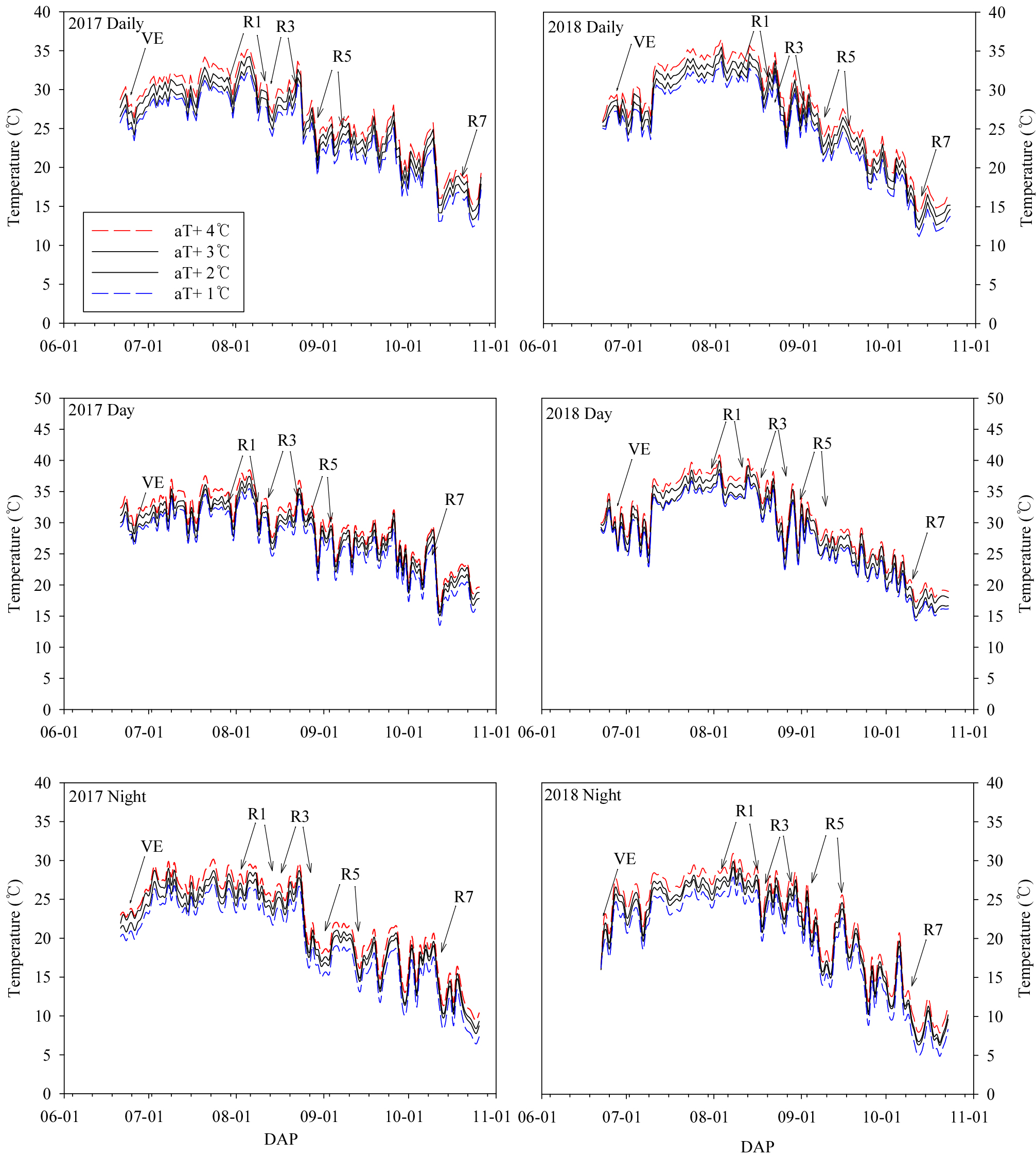
2017년과 2018년 대원콩, 풍산나물콩 및 대풍콩은 온도구배에 따라 생물계절이 차이를 보였다(Table 1). 2017년 대원콩과 대풍콩은 온도구배에 따라 차이 없이 7월 25에서 7월 27일에 개화하였고, 풍산나물콩은 7월 31일에서 8월 1일에 개화하였다. 착협기는 대원콩이 8월 13일에서 14일이고, 풍산나물콩은 8월 20일에서 21일이었고, 대풍콩은 8월 16일에서 19일에 이루어졌다. 종실비대기는 대원콩이 8월 26일에서 8월 30일이고, 풍산나물콩은 9월 1일에서 9월 4일이었고, 대풍콩은 8월 25일에서 8월 28일에 이루어졌다. 종실비대성기에서는 대원콩과 풍산나물콩 및 대풍콩의 aT+ 4가 다른 온도구배에 비하여 각각 5일에서 6일과 4일 그리고 4일이 지연되었다.
Table 1. Effect of elevated temperature on duration of soybean growing stages during 2017 and 2018.
yAmbient temperature+ 1°C: aT+ 1, ambient temperature+ 2°C: aT+ 2, ambient temperature+ 3°C; aT+ 3, ambient temperature+ 4°C: aT+ 4
xBeginning flowering: R1, beginning pod: R3, beginning seed: R5, beginning maturity: R7.
Day of year: Doy
2018년 개화기는 대원콩이 온도구배에 따라 7월 28일에서8월 6일로서 특히 aT+1에 비하여 aT+4가 9일이 지연되었다. 풍산나물콩은 8월 6일에서 8월 9일에 이루어, 대풍콩은 일률적으로 7월 28일에 개화하였다. 착협기는 대원콩, 풍산나물콩 및 대풍콩이 aT+1에 비하여 aT+4가 약 3-6일이 지연되었다. 종실비대기는 대풍콩이 온도구배에따라 뚜렷하게 지연되었는데 aT+1에 비하여 aT+4가 9일이 지연되었다. 풍산나물콩은 aT+1에 비하여 aT+4가 6일이고 대풍콩은 aT+1에 비하여 aT+4가 4일이 지연되었다. 종실비대성기는 대원콩, 풍산나물콩 및 대풍은 aT+3과 aT+4는 aT+1과 aT+2에 비하여 3-6일이 지연되었다.
수량구성요소와 수량 변화
2017년과 2018년, 품종 및 온도구배에 따라 주경 마디수와 수량 구성요소 및 종실 수량은 고도의 유의성을 보였다(Table 2). 2017년과 2018년 수량 구성요소는 고도로 유의하였다. 특히 2017년에 비하여 2018년이 수량 구성요소 및 종실 수량이 급격히 낮았다. 품종간 수량 구성요소는 고도의 유의성을 보였다. 온도구배간에서 종실 수량을 제외한 수량 구성요소는 유의성을 보였다. 연차 × 품종간에는 협 당 립수를 제외한 수량구성 요소는 고도의 유의성을 보였다. 반면, 연차 × 온도구배에서는 협 당 립수와 종실 수량은 유의성이 크지 않았다. 그러나 품종 × 온도구배간에는 수량 구성요소와 종실 수량은 고도의 유의성을 보였다. 연차 × 품종 × 온도구배에는 협 당 립수를 제외한 다른 수량 구성요소와 종실 수량은 고도로 유의하였다.
Table 2. Analysis of variance to determine the effect of year, cultivar, temperature, and interactions among them on soybean node number, yield components, and seed yield.
zDaewonkong: DWK, Pungsannamulkong: PSNK, Deapongkong: DPK
yAmbient temperature+ 1°C: aT+ 1, ambient temperature+ 2°C: aT+ 2, ambient temperature+ 3°C; aT+ 3, ambient temperature+ 4°C: aT+ 4
품종에 따른 온도구배는 고도의 유의성을 보였다(Table 3). 2017년 대원콩은 aT+4에서 면적당 협수와 100립 중이 높아 종실 수량이 높았다. 풍산나물콩은 aT+4가 다른 온도구배에 비하여 100립 중이 낮아서 종실 수량이 낮았다. 대풍콩은 협수 당 종실, 100립중 및 종실 수량이 고도로 유의하였다. 100립 중은 aT+4가 aT+2와 aT+3에 비하여 낮았다. 2018년은 품종과 온도구배간에는 유의한 차이를 보였다. 대원콩과 풍산나물콩은 면적당 협수, 협 당 립수 및 100립 중은 aT+4가 다른 온도구배에 비하여 낮아서 종실 수량도 낮았다. 대풍콩은 온도구배간 100립 중을 제외한 수량 구성요소는 유의한 차이가 나지 않았다.
Table 3. Effect of elevated temperature on soybean node number, yield components, and seed yield.
zDaewonkong: DWK, Pungsannamulkong: PSNK, Deapongkong: DPK
yAmbient temperature+ 1°C: aT+ 1, ambient temperature+ 2°C: aT+ 2, ambient temperature+ 3°C; aT+ 3, ambient temperature+ 4°C: aT+ 4
온도 상승과 종실 단백질 및 지방함량 변화
온도 상승에 따른 콩 종실의 단백질과 지방 함량 변화는 연차, 온도구배 및 품종간 고도의 유의성을 보였다(Table 4). 단백질과 지방함량은 2017년에 비하여 2018년이 각각 11%와 3.6%가 감소하였다. 연차간과 품종간에는 단백질과 지방함량은 고도의 유의성을 보였다. 그러나 온도구배간 지방함량은 유의성이 나타나지 않았다. 연차 × 품종, 연차 × 온도구배, 품종 × 온도구배 및 연차 × 품종 × 온도구배에서는 단백질과 지방함량은 고도의 유의성을 보였다.
Table 4. Analysis of variance to determine the effect of year, cultivar, temperature, and interactions among them on the protein and oil content of soybean seeds at R8.
zDaewonkong: DWK, Pungsannamulkong: PSNK, Deapongkong: DPK
yAmbient temperature+ 1°C: aT+ 1, ambient temperature+ 2°C: aT+ 2, ambient temperature+ 3°C; aT+ 3, ambient temperature+ 4°C: aT+ 4
2017년과 2018년 대원콩, 풍산나물콩 및 대풍콩의 온도구배에 따라 단백질과 지방함량은 유의성을 보였다(Table 5). 2017년 대원콩과 풍산나물콩은 온도 상승에 따라 단백질과 지방함량은 각각 감소와 증가를 하였다. 그러나 대풍콩은 온도 상승에 따라 단백질 함량은 aT+3에서 높았으나 aT+4에서 감소하였고 지방함량은 온도구배에 따라 유의한 차이가 나타나지 않았다. 2018년 대원콩의 단백질과 지방함량은 aT+3에서 가장 높았고 aT+4에서 감소하였다. 풍산나물콩은 온도 상승에 따라 단백질함량은 증가하였지만 지방함량은 감소하였다. 대풍콩은 온도 상승에 따라 단백질함량이 증가하고 지방함량은 감소하였다.
Table 5. Effect of elevated temperature on soybean seed protein and oil content at R8.
zDaewonkong: DWK, Pungsannamulkong: PSNK, Deapongkong: DPK
yAmbient temperature+ 1°C: aT+ 1, ambient temperature+ 2°C: aT+ 2, ambient temperature+ 3°C; aT+ 3, ambient temperature+ 4°C: aT+ 4
고 찰
생물계절 및 착협수 변화
본 연구는 재배기간 온도상승에 따라 콩의 생물 계절, 수량구성소요, 단백질 및 지방함량 변화에 대한 결과이다. 일반적으로 평균온도가 30°C 이상을 넘으면 개화기와 종실비대기가 지연된다 하였다(Dornbos & Mullen, 1991; Gibson & Mullen, 1996a; Zheng et al., 2002; Egli et al., 2005). 본 연구에서 대원콩과 풍산나물콩의 개화기는 aT+1에 비하여 aT+3과 aT+4°C가 3일에서 9일이 지연되었다. 반면 대풍콩은 온도구배와 관계없이 일률적으로 개화기와 착협기가 진행하였다. 종실비대기에는 대원콩, 풍산나물콩 및 대풍콩이 aT+3과 aT+4°C가 aT+1에 비하여 3일에서 9일이 지연되었다(Table 1).
온도구배에 따른 면적당 협수는 유의한 차이가 크지 않았다. 2017년에 비하여 2018년의 대원콩과 풍산나물콩의 협수는 심각하게 감소하였다. 반면 대풍콩은 2017년과 2018년은 비슷한 협수를 확보하였다. 이는 착협시기인 8월 중순에 평균 온도가 30°C 이하로 내려가면서 고온 피해를 피한 것으로 판단된다. 반면, 대원콩과 풍산나물콩은 개화기의 지연으로 착협기가 다소 지연되면서 30°C이상의 고온 피해를 받은 것으로 판단되었다(Table 1 and Table 2).
개화기에서 착협기까지 주야간 온도가 30/22°C에서 개화률과 결협률이 증가를 하고, 착협기간 동안 주야간 온도가 26/20°C에서는 개체당 협수가 증가하지만(Dornbos & Mullen, 1991; Gibson & Mullen, 1996a; Zheng et al., 2002; Egli et al., 2005), 이 범위를 벗어 나게 되면 결협률이 떨어진다고 하였다(Djanaguiraman et al., 2013). 개화성기에서 착협시 사이의 평균 온도 30°C 이상을 벗어나게 되면 수술의 개폐가 실패 되고, 암술로 관통하는 화분이 감소하여 결협률이 감소한다고 하였다(Djanaguriaman et al., 2013; Allen et al., 2016; Sita et al., 2017). 또한 낮 기온 27-33°C와 밤 기온 19-24°C에서는 착협수가 감소한다고 하였다(Thuzar et al., 2010).
종실 크기와 수량 변화
등숙기간의 고온은 크기를 억제한다 하였다(Sato & Ikeda, 1979; Matsuda et al., 2011; Choi et al., 2016). 본 연구결과 풍산나물콩과 대풍콩의 aT+4는 aT+1에 비하여 등숙기간 단축과 100립 중이 감소하였다. 특히, 2018년은 심각한 고온으로 등숙기간 단축과 함께 자엽의 크기가 정상적으로 도달하지 못 하여 100립 중이 감소한 것으로 판단되었다(Table 3).
등숙속도와 관련하여 주야간 온도가 32/22°C에서 가장 활발하고, 32/22°C를 넘으면 지연 현상을 보이다가 36/26°C에서는 심각하게 지연된다 하였다(Dornbos & Mullen, 1991). 종실 크기와 관련하여 주간 온도 30-35°C와 야간 온도 20- 25°C에서 15-18%의 종실 무게가 감소한다 하였다(Gibson & Mullen, 1996a). 30°C이상의 고온이 지속될수록 종실로 전류 하는 동화물질량과 기간이 단축되어 종실 크기가 급격히 감소한다 하였다(Puteh et al., 2013). 종실비율과 최종 종자 크기는 세포 수와 세포 크기 그리고 동화물질의 공급능력에 의해 결정된다 하였다(Djanaguiraman et al., 2013). 등숙기간 온도와 관련하여서 주야간 온도가 28/18°C와 40/30°C에서는 상승된 온도의 종자 크기, 속도 지연과 함께 등숙기간 단축되어서 자엽 당 세포수가 감소한다고 하였다(Thomas et al., 2010). 고온은 자엽 세포 확대를 제한하여 종실 크기를 감소시켜 약 38-40%의 종실 수량이 감소한다 하였다(Tacarindua et al., 2012; Tacarindua et al., 2013). 본 연구결과 고온의 피해가 심각하였던 2018년이 2017년에 비하여 100립 중이 약 27%가 감소하였다. Uchikawa et al. (2003)은 1985년-2000년 작황 자료를 분석한 결과 개화기에서 종실비대기까지의 평균온도와 100립중을 비교한 결과 평균온도 25°C이상부터 100립 중이 감소한다 하였다. Matsuda et al. (2011)은 1989-2008년 작황 자료를 분석한 결과 개화기에서 성숙기의 최고 온도 27°C 이상과 평균 온도 22°C이상 그리고 최저 기온 18°C 이하에서는 100립 중이 감소한다 하였다.
Zheng et al. (2009)은 20년(1987-2007년) 간 포장 자료를 기반으로 종실비대기(R5)-등숙기(R7) 단계에서 최대 일일 평균 온도 20-24°C에서 6-10%의 수량이 증가 하였다. 그러나 등숙기간 주간 최고 온도 29-34°C와 야간 평균온도 20°C에서는 수량에 지장을 준다 하였다(Dornbon & Mullen, 1991). 또한 Egli & Wardlaw (1980)에 의하면 등숙기간 주야간 온도가 24/19°C보다는 33/28°C에서 종실 수량이 약 22%가 감소한다 하였다. 따라서 평균온도 30°C이상을 넘으면 결협률, 착협수 및 종실비대 속도 지장과 함께 수량 저해를 초래한다 하였다(Mochizuki et al., 2005). 본 연구결과 고온인 2018년이 2017년에 비하여 수량이 54%가 감소하였다. 그러나 온도구배간에는 뚜렷한 차이는 보이지 않았다(Table 2).
종실 단백질과 지방함량 변화
재배기간 고온은 소스의 잠재적 능력과 싱크의 저장능력을 불 균일하게 일어나는데, 특히, 등숙기간 고온 스트레스는 동화 물질을 저장 기관으로 분배하는 과정에서 불 균일하게 진행이 된다 하였다(Egli et al., 2005). 본 연구에서 대원콩, 풍산나물콩 및 대풍콩은 온도구배에따라 단백질과 지방함량이 감소 또는 증가하였다. 2018년은 2017년에비하여 대원콩과 풍산나물콩은 단백질과 지방함량이 급격하게 감소하였다. 그러나 대풍콩의 2018년은 2017년에 비하여 지방함량이 높았다. 이러한 결과는 개화 후 3주 평균온도가 2017년에 비하여 2018년이 약 1°C가 낮아졌기 때문에 지방함량이 높은 것으로 판단된다(Table 4 and Fig. 1).
콩은 개화 후 3주 정도가 되면 종실이 비대해지면서 동화물질이 종실로 전류를 시작하는 시기이다(Poeta et al., 2016). 일반적으로 성숙 종실의 지방함량의 50%는 개화 후 3-4주 정도에 축적이 되고, 단백질함량은 개화 후 4-5주 정도부터 성숙 종실에 50%가 축적이 된다 하였다(Yazdi et al., 1977; Wilson et al., 1978). Rotundo&Westgate (2009)에 의하면 환경 요인과 관련하여 단백질과 지방함량을 분석한 결과 단백질 함량은 고온으로 약 9%이상이 감소하고, 지방함량은 15% 이상 감소한다 하였다. 또한 성숙군과 연차에 따라 단백질과 지방함량에 영향을 준다 하였다(Bellaloui et al., 2011). 재배기간 고온은 잎의 질소함량을 감소시키고 호흡량을 증가시켜서 동화물질의 공급을 억제한다 하였다(Gibson&Mullen, 1996b; Dornbos&Mullen, 1992; Thomas et al., 2003; Rotundo et al., 2009; Xu et al., 2016). 생식 생장 기간 동안 평균 온도 22-36°C에서는 단백질함량이 증가를 하지만 이 범위를 벗어나게 되면 감소한다 하였다(Piper&Boote, 1999; Bellaloui et al., 2016; Xu et al., 2016).
고온으로 인한 지방함량 감소는 개화기에서 종실비대기사이에는 영향이 작지만, 종실비대기에서 수확기사이에는 20% 이상 감소한다 하였다(Rotundo&Westgate, 2009). 지방함량은 평균온도 25-28°C에서는 증가하고 이 범위를 넘으면 감소한다 하였다(Dornbos&Mullen, 1992; Gibson&Mullen, 1996b; Piper&Boote, 1999; Bellaloui et al., 2016; Xu et al., 2016; Mourtzinis et al., 2017). Song et al. (2016)은 2010년부터 2013년까지 763개의 콩 종실에 대한 단백질과 지방함량을 분석한 결과 단백질함량은 누적 온도, 일 평균 온도, 강수량에 긍정적인 영향을 주지만, 지방함량은 일조 시간과 일일 누적온도에는 부정적인 영향을 준다 하였다. 본 연구 결과 온도구배에 따라 일부 콩의 주요 형질적 특성은 변화하였다. 그러나 이러한 형질적 특성과 온도구배챔버와 관련하여 결과를 도출하기에는 다소 한계의 소지를 부여하였다.
적 요
본 연구는 자연 환경과 가장 유사한 온도구배 챔버에서 국내 최대 보급 품종인 대원콩, 풍산나물콩 및 대풍콩을 통해 재배기간 온도 상승에 따른 생물 계절, 수량구성요소, 단백질 및 지방함량 변화를 구명하고자 수행을 하였다.
2017년과 2018년 대원콩과 풍산나물콩은 aT+ 1에 비하여 aT+ 4에서 개화기가 지연하였다. 특히 2018년이 지연 일수가 길었다. 반면 대풍콩은 일률적으로 개화를 하였다. 이러한 결과 등숙기간은 년차간과 온도구배간 약 3-9일이 지연이 되었다. 수량 구성요소에서는 2017년에 비하여 2018년이 감소 폭이 높았다. 특히 100립 중과 종실 수량이 감소하였다. 연차와 품종에 따라서 면적 당 협수의 감소는 대원콩과 대품콩이 각각 48.8%와 41.5%씩 감소를 하였다. 풍산나물콩은 14.7%가 감소를 하였다. 단백질과 지방함량은 연차, 품종 및 온도구배에 따라 고도의 유의성을 보였다. 특히 2018년이 2017년에 비하여 등숙기간 온도 상승으로 인해 단백질과 지방함량이 감소하였다. 그러나 대풍콩은 2017년에 비하여 2018년이 지방함량은 높았다.
본 연구에 알 수 있듯이 개화기에서 종실비대기사이의 온도 상승은 생물 계절을 지연과 협수와 100립 중이 감소하여 종실 수량 감소로 이어졌다. 또한 콩 종실의 주요 성분인 단백질과 지방함량을 감소시켰다. 따라서 향후 지구온난화로 인한 수량 보다는 영양학적 측면도 연구가 진행되어야 할 것으로 판단된다.
