

서 론
재료 및 방법
시험재료 및 강우 처리
완전미율, 단백질 함량, 전분호화특성
통계처리
결 과
고 찰
등숙기 기상환경 조건에 따른 수발아 발생 변이
등숙기 온도 및 수발아 발생에 따른 품질변이
적 요
서 론
벼는(Oryza sativa L.) 전 세계 인구의 절반 이상이 주식으로 하는 세계에서 가장 중요한 식량작물중 하나이며, 인간이 섭취하는 최종 수확물인 벼 종실의 수량과 품질은 수확 전 등숙기 기상환경에 크게 좌우된다(Lin et al., 2011; Zhu et al., 2019). 최근 지구온난화로 인한 일시적 고온 및 등숙기 평균기온의 증가, 게릴라성 집중호우, 태풍과 같은 이상기상현상 발생 빈도 증가로 종실의 수량 및 품질을 저하시키는 수발아 피해 위협이 증가하고 있다(Choi et al., 2000; Lee et al., 2006; Falade et al., 2014).
수발아는 벼가 수확 전 포장에서 발아가 가능한 일정한 온도와 수분조건에 노출되어 종실이 이삭에 달려있는 상태로 발아 하는 것을 의미한다. 발아를 위한 외적 환경조건이 갖추어진 상태에서는 유아를 생장시키기 위해 종실 주요 구성성분인 전분, 단백질들을 분해하여 에너지를 생성한다(Vetch et al., 2019). 이러한 과정에는 종피 투수성과 같은 물리적 특성, a-amylase 등의 효소 활성 유도, ethylene, ABA, GA 등의 호르몬 함량, 종자의 휴면성과 같은 다양한 내적요인들이 관여되어 있다(He & Yang, 2013; Lee et al., 2016).
이러한 요인들의 변이는 종실의 등숙 단계에 따라서 발생되어 수발아 발생에 영향을 미친다. 등숙 단계별로 수발아 발생 양상을 보면 등숙 후기인 황숙기 이후에 발생률이 높아진다고 하는데(Park & Park, 1984), 이는 종실이 발달되면서 일정 시점에 ABA 함량이 최대가 되었다가 이후 성숙까지 점차 감소하는 특성(Seo & Koshiba, 2002; Zhang et al., 2012)과 종실이 출수 후 일정한 적산온도를 경과한 이후부터 발아가 가능해지는 특성(Baek & Chung, 2014)이 영향을 미치는 것으로 생각된다. 이러한 특성은 등숙기 기상환경, 특히 평균기온의 변화에 따라서 출수 후 동일한 일수가 경과함에도 불구하고 크게 달라진다(Hakata et al., 2012; Kang et al., 2018).
한편 수발아가 발생하면 종실의 품질 변이에 영향을 미친다는 결과들이 보고된 바 있다. 수발아 발생 시 유아 길이에 따라서 완전미율 및 도정률이 감소하였고(Kim & Lee, 1996; Oh et al., 1987), 전분호화 특성이 감소하지만, 단백질함량에는 변화가 없었다고 보고되었다(Kim et al., 2008). 최근에는 수발아 발생에 따라 전분구조의 변화를 분석하여 호화특성 및 단백질 함량 등 이화학적 특성과의 관계를 분석한 내용이 보고되었다(Zhu et al., 2019). 하지만 수발아 발생에 따른 전분호화특성과 단백질 함량 등의 이화학적 특성 변이를 비교한 결과는 수발아 발생 전, 후 조건으로만 비교 분석되어, 수발아 발생 정도 별 변화 추이를 확인하기에는 미흡한 부분이 있다.
본 시험에서는 이앙시기를 조절하여 등숙 특성에 영향을 미치는 큰 요인 중 하나인 평균기온 변이에 따른 수발아 발생 정도를 분석하고, 각 등숙온도 조건에서 수발아 발생 정도 별 종실의 품질 및 이화학적 특성 변이에 대한 결과를 분석하였다.
재료 및 방법
시험재료 및 강우 처리
본 시험에서는 출수 생태형별로 2품종씩 조생종은 오대, 조평, 중생종은 대보, 하이아미, 중만생종은 새누리, 신동진을 시험 재료로 이용하였다. 2018년과 2019년 모두 육묘상자에서 20일동안 육묘한 후 1차 시기는 조생종은 6월 6일, 중생·중만생종은 5월 16일, 2차 시기는 모든 생태형 및 품종 동일하게 6월 26일에 이앙하였다. 1차 이앙 시기를 달리한 이유는 등숙기 온도 차이를 유사하게 설정하기 위해 비교적 비슷한 시기에 출수할 수 있도록 조생종은 중생, 중만생종보다 늦은 6월 6에 이앙하였다. 비료는 N-P2O5-K2O: 9-4.5-5.7 (kg/10a)을 기준으로 시비하였으며, 그 밖의 재배관리는 국립식량과학원의 벼 표준재배법에 준하여 재배하였다. 수발아 처리를 위한 재료는 각 이앙시기 및 생태형 품종별 출수 후 적산온도 1200°C에 도달하는 날을 기준으로 포장에서 뿌리 위 바로 위쪽에서 절단하여, Fig. 1에서 보는 바와 같이 이삭이 줄기에 달려있는 상태로 국립식량과학원 인공기상시설에서 강우 강도 5.2 mm/hr 조건에서 조생종은 평균온도 24°C (최고 29°C/최저 19°C), 중생·중만생종은 평균온도 22°C (최고 27°C/최저 17°C)으로 0, 24, 48, 72, 96, 120, 144시간까지 처리하였다. 각 처리 별 수발아율은 30개 이삭을 분석하여 평균하였다. 수발아율은 수발아 처리 후 종실의 수분이 마르게 되면 확인이 어려워질 수 있기 때문에 강우처리 후 즉시 유아가 왕겨를 뚫고 나온 것을 확인하였다. 왕겨가 벌려져 있는 상태이지만 유아의 확인이 애매한 경우의 종실은 왕겨를 벗겨 유아의 존재를 확인하였다. 생태형 및 품종 간의 수발아율 발생 차이를 비교하는 것이 아닌 각 생태형 및 품종별 등숙기 평균기온에 따른 수발아 발생 변이를 확인하는 실험이기 때문에, 강우 처리 시 온도는 생태형별로 실제 포장에서 수발아가 발생할 수 있는 시기의 강우 온도를 설정하기 위하여 출수 후 적산온도 800°C~1200°C 시기의 평균기온을 고려하여 조생종은 24°C, 중생·중만생종은 평균온도 22°C로 설정하였다.

완전미율, 단백질 함량, 전분호화특성
완전미율 등의 외관품위는 Rice Quality Analyzer (RN300 model, Kett, Japan)을 이용하여 측정하였다. 단백질함량의 분석은 분쇄한 시료(종실) 250 mg을 Elementar Analyzer System (Vario Macro, Germany)을 이용하여 분석한 총 질소함량 값과 켈달장치를 이용하여 분석한 값을 비교하여 보정계수를 구하고 보정계수를 각 분석 값에 곱하여서 구하였다. 호화특성은 Rapid-Visco Analyser (Newport Scientific Model RVA-4, Australia)를 이용하여 AACC Method 61-02 에 의하여 peak viscosity (최고점도), trough viscosity (최저점도), breakdown (강하점도), final viscosity (최종점도)를 측정하였다.
통계처리
각 실험결과의 통계처리는 R 통계프로그램(Version 3.2.2)를 이용하여 T-검정 및 분산분석(ANOVA)후 Duncan’s multiple range test로 p < 0.05 수준에서 유의적 차이를 검정하였다. piecewise regression (segmented regression) 분석은 sigmaplot 11.1을 이용하여 분석을 진행하였다. Breaking point x = c일 때 아래의 piecewise regression 모델이 사용되었다(Ryan & Porth, 2007).
결 과
2018년과 2019년의 각 생태형 및 품종별 이앙시기에 따른 출수기와 출수 후 40일간 등숙기 평균기온은 Table 1에 나타낸 바와 같다. 각 품종별 이앙시기에 따라서 출수기의 변동으로 6월 26일에 이앙한 처리구 대비 5월 16일, 6월 6일에 이앙한 처리구에서 등숙기 평균기온이 3.1°C에서 3.7°C까지 증가한 결과를 보였다.
Table 1.
Changes in heading date and average temperature during the ripening stage according to transplanting dates in 2018 and 2019.
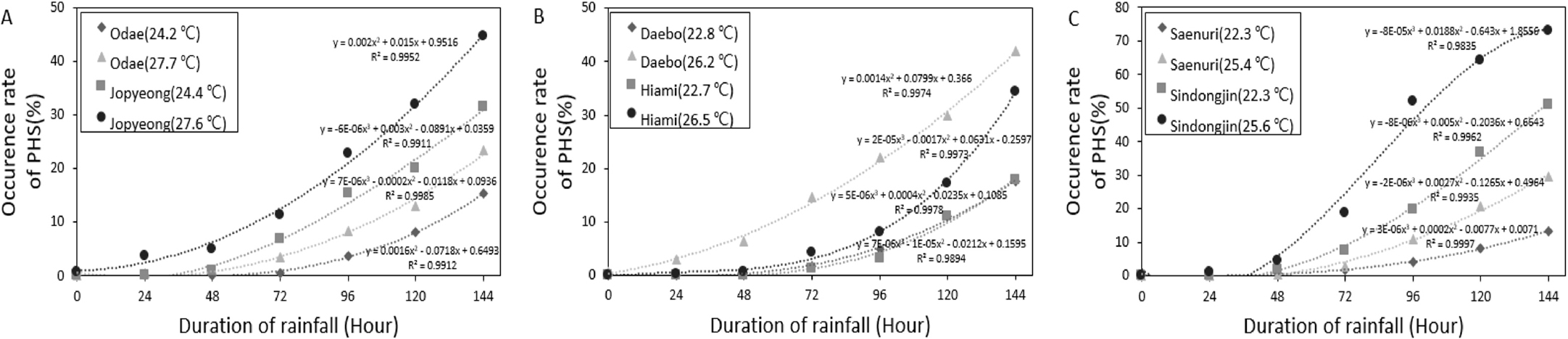
이러한 등숙기 온도 변이에 따른 수발아 발생율의 차이는 Fig. 2에 나타낸 바와 같다. 출수 후 적산온도 1200°C시점에서 동일한 강우조건으로 144시간까지 처리했음에도 불구하고 등숙기 평균기온이 상대적으로 고온이었던 처리구에서 오대의 경우 8%, 조평은 13%, 대보는 24%, 하이아미는 17%, 새누리는 16%, 신동진은 22%까지 모든 품종에서 수발아 발생률이 증가한 것을 확인 할 수 있었다.
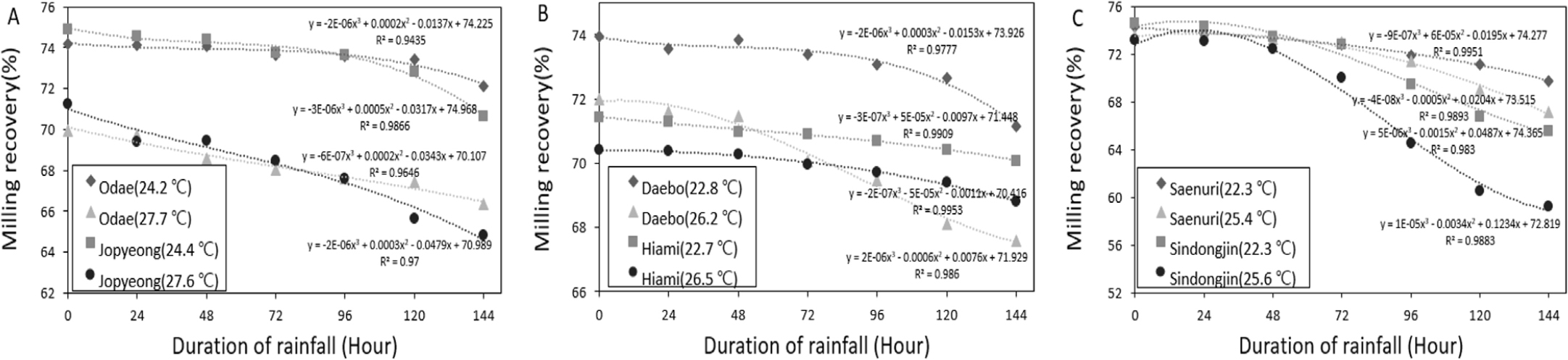
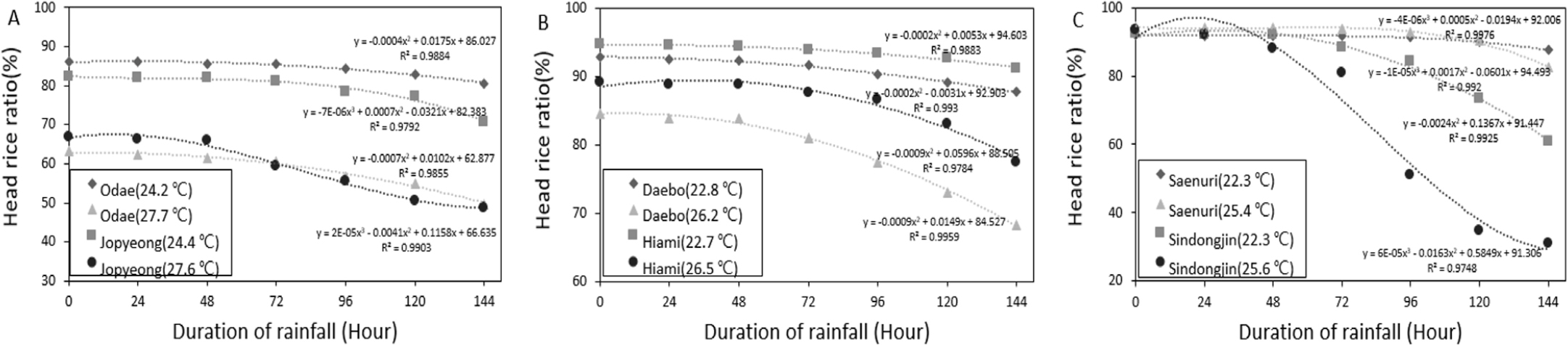
강우가 지속될수록 수발아율이 증가함에 따라서 도정률 및 완전미율이 감소하였던 결과는 Figs. 3, 4에 나타낸 바와 같다. 먼저 강우를 처리하지 않은 조건에서 온도 별로 비교하였을 때 상대적으로 고온처리구에서 도정률 및 완전미율 자체가 낮았으며, 강우처리에 따라서는 수발아율이 증가하였을때 도정률 및 완전미율 감소 정도가 고온처리구에서 적온처리구 대비 컸던 결과를 보였다. 결국 동일한 강우조건에서도 등숙기 고온조건에서 도정률 및 완전미율이 훨씬 낮아 품질이 불량하게 되었던 결과를 확인할 수 있었다.
등숙온도 및 수발아 발생 정도별 완전미율 감소에 따른 분상질립, 싸라기 등 기타 외관품위 변이 정도를 나타낸 결과는 Fig. 5에 나타낸 바와 같다. 지속강우가 처리되지 않은 조건에서는 상대적으로 고온처리구에서 적온처리구 대비 싸라기가 약 7%, 분상질립이 1% 높아 완전미율이 감소하였으며, 강우처리에 따른 수발아율 증가 시 분상질립 및 싸라기의 증가로 완전미율이 감소하는 것을 확인 할 수 있었다. 또한 등숙기 적온 처리구에서 강우처리 144시간 노출로 수발아가 24% 발생했던 종실에서는, 강우 처리전 대비 싸라기가 8% 증가되었는데, 고온 처리구에서는 96시간 노출에 따라서 수발아가 21%로 적온 처리구 대비 약 3% 더 낮게 발생 되었음에도 강우 처리전 대비 싸라기가 11% 증가되어, 고온처리구에서는 더 낮은 수발아 피해가 발생했음에도 불구하고 외관품위 감소율이 더 컸던 결과를 확인 할 수 있었다.
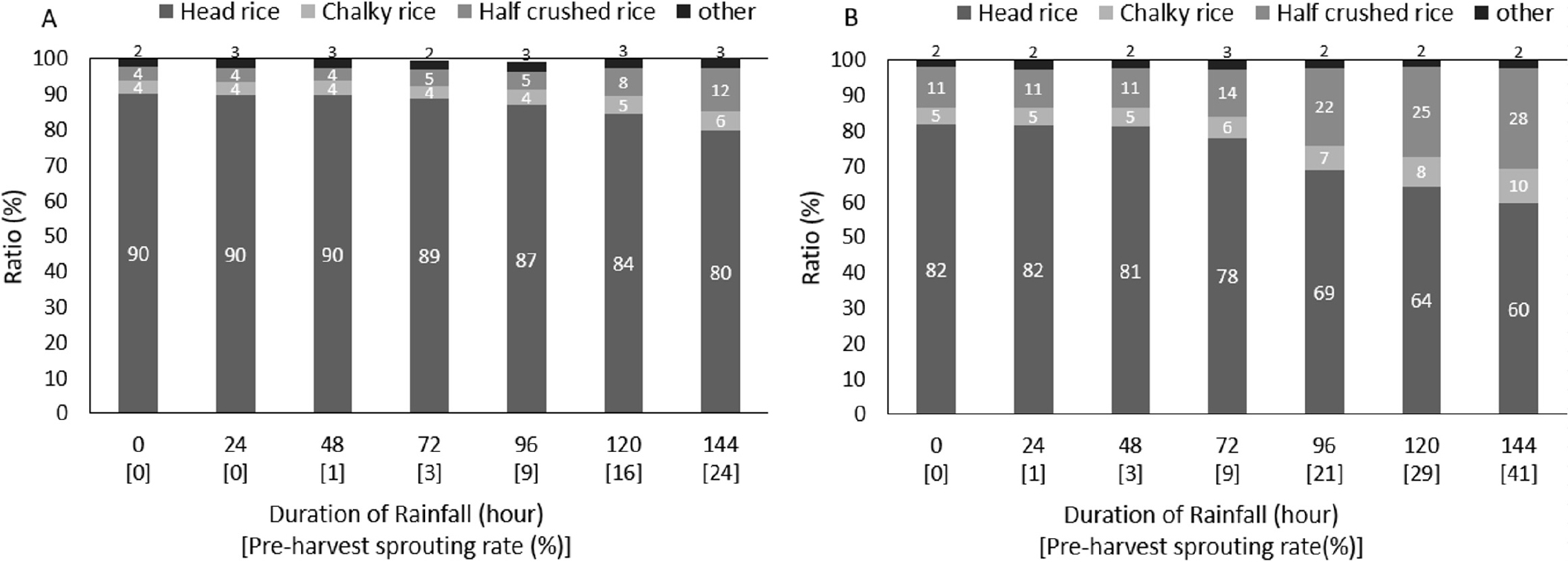
Fig. 5
Changes in the proportion of each appearance grain quality (average of six varieties) by rainfall treatment and average temperature during the ripening period. A: 23.2°C (optimum temperature treatment in the ripening period for the average of six varieties). B: 26.3°C (high temperature treatment in the ripening period for the average of six varieties).
등숙기 온도 및 수발아 발생 정도에 따른 전분호화특성 변이를 나타낸 결과는 Fig. 6에 나타낸 바와 같다. 최고, 최저, 최종점도는 종실에 열과 수분을 가하였을 때 호화되어 부풀어 오르는 특성과 관련되며, 강하점도는 최고점도와 최저점도간의 차이로 전분입자가 열에 의해 붕괴되는 정도를 나타낸다. 치반(Setback)점도는 최종점도와 최저점도간의 차이로 노화 특성을 나타낸다(Ha et al., 2006; Chun, 2009). 지속강우가 처리되지 않은 종실에서는 등숙기 고온조건에서는 적온조건 대비 최고, 최저, 최종점도 및 강하점도가 증가하였으며, 치반점도가 감소한 결과를 보였다. 강우처리에 따라서 두 등숙기온도 조건에서 모두 수발아율 1% 까지는 최고점도가 다소 증가하였다가 그 이후부터는 감소하는 경향을 보였다.
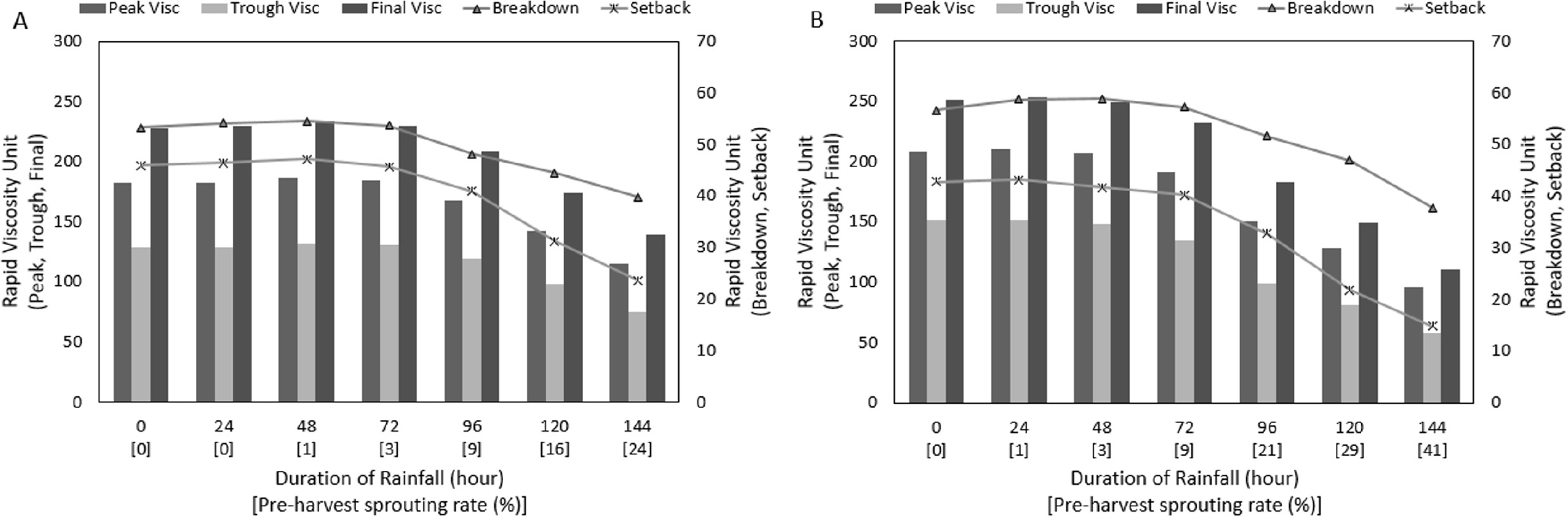
Fig. 6
Changes in starch gelatinization of rice grains by rainfall treatment and average temperature during the ripening period. y-axis: peak, trough, and final viscosity (left); breakdown and setback (right). Visc: viscosity. A: 23.2°C (optimum temperature treatment in the ripening period for the average of six varieties). B: 26.3°C (high temperature treatment in the ripening period for the average of six varieties).
등숙기 온도 및 품종 별 수발아율에 따른 단백질함량 변이를 나타낸 결과는 Fig. 7에 나타낸 바와 같다. BreakPoint는 piecewise 회귀분석에 의하여 결정되는 계수로 회귀선의 기울기가 변이되는 시점을 나타낸다. 온도에 따라서는 오대의 경우 등숙기 고온조건에서 단백질함량이 감소하는 시점(BreakPoint)이 적온 조건 대비 수발아율이 높은 조건이었지만, 그외 모든 품종에서는 등숙기 고온조건에서 감소 시점이 수발아율이 더 낮은 조건이었던 것을 확인 할 수 있었다. 또한 온도조건이 높을수록 수발아 발생율 증감에 따른 단백질 함량의 변이가 다소 컸던 결과를 확인할 수 있었다.
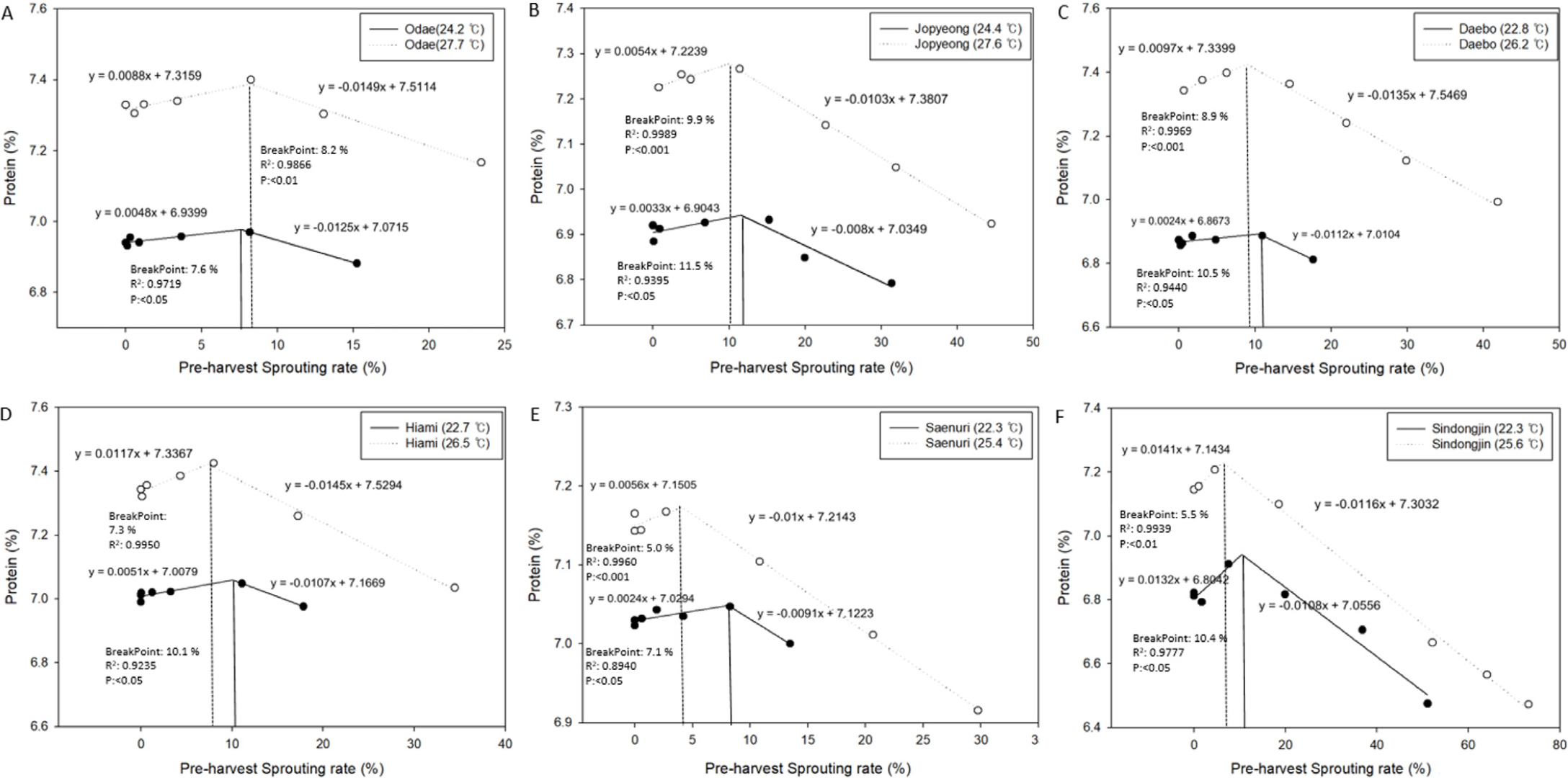
Fig. 7
Piecewise (segmented) regression analysis between the preharvest sprouting rate and protein content for each cultivar. A: Odae, B: Jopyeong, C: Daebo, D: Hiami, E: Saenuri, F: Sindongjin. Only insignificant changes in protein content occurred up to the break point in each case, followed by a rapid fall after the break point (preharvest sprouting rate).
고 찰
등숙기 기상환경 조건에 따른 수발아 발생 변이
수발아 저항성을 높인 품종을 개발하기 위한 수발아 발생 메커니즘, QTL, Key gene을 밝히는 등의 다양한 연구들이 진행되고 있다(Masojc & Milczarski, 2008; Nonogaki & Nonogaki, 2017). 보통 수발아 내성 검정을 위한 실험을 진행할 때 등숙 단계별로 ABA, GA 함량, a-amylase 등의 수발아와 관련된 요인들이 달라지는 것을 인지하고 이에 대한 변이를 줄이기 위하여 대부분 출수, 개화 후 30 일, 40 일 등 특정 일수에 맞춰서 수발아 내성을 검정한다(Gao et al., 2008; Park & Kim, 2009). 하지만 출수, 개화 후 특정 일수는 출수, 개화 후 등숙기 평균기온에 의하여 크게 차이 난다. 예를 들어 출수 후 적산온도 1200°C까지 도달하는 출수 후 일수는 등숙기 평균기온이 22°C의 온도에서는 55 일, 25°C에서는 약 48 일, 28°C에서는 약 43 일이 되어 동일한 출수 후 일수임에도 고온조건에서는 상대적으로 등숙후기에 처리가 이루어지게 된다. 결국 수발아율 변이에 영향을 미칠 수 있는 ABA 함량, 전분립의 구조 등의 요인들이 출수 후 일수에 따라서 고정되지 않고 변동이 생기며(Liu et al., 2014; Vetch et al., 2019), 동일한 품종 내 온도에 따른 수발아율 변이가 품종 간 차이보다 커지는 결과를 얻을 수 있는 상황이 발생할 가능성이 생기게 된다(Fig. 2). 결국 수발아 검정을 위해서는 수발아를 처리하는 시점의 온도, 수분 처리조건 등도 중요하지만 처리시점에 따른 변이가 크게 작용 할 수 있기 때문에, 출수 후 일수가 아닌 적산온도를 기준으로 조건을 맞춰 시험을 진행해야 한다. 한편 고온등숙시 수발아 발생율이 증가하기 때문에(Fig. 2) 등숙기 온도조건을 고려하지 않고 이앙 시기를 무조건 빨리 앞당기게 된다면 수발아 발생 위험을 증가시킬 수 있기 때문에 등숙기 온도조건을 고려하여 이앙시기를 설정해야 한다.
등숙기 온도 및 수발아 발생에 따른 품질변이
본 시험에서는 등숙기 온도별 수발아 발생정도에 따른 완전미율, 도정율 뿐만 아니라 이에 따른 종실의 이화학적 특성 변이 결과를 함께 연결하여 분석하였다. 종실이 고온 등숙되는 조건에서는 상대적으로 전분을 합성하는 효소의 활성은 증가하지만, 전분합성에 필요한 종실에 공급되는 동화산물의 양은 그 수요를 맞추지 못하고 지속기간이 짧아져 등숙에 필요한 충분한 기간이 주어지지 못하게 된다(Lee et al., 2017; Yang et al., 2007). 결국 전분립의 입자가 작아지고 전분이 불완전하게 채워진 형태의 종실이 되어 무게 및 전분함량이 감소하며, 단백질함량, 분상질립 등이 증가하여 품질이 저하된다(Lisle et al., 2000; Tashiro & Wardlaw, 1991). 본 시험에서도 마찬가지로 상대적으로 등숙기에 고온인 조건에서 립중, 완전미율, 도정률이 감소하고, 단백질함량, 분상질립 등이 증가하였던 결과를 확인하였다. 또한 수발아율 증가에 따라서 완전미율이 감소하였으며, 이는 싸라기 및 분상질립의 증가 때문이었다(Fig. 5). 이러한 분상질립의 증가는 단순 외관품이의 저하뿐만 아니라 이화학적 특성 변화에 크게 영향을 미친다. 수발아 발생한 벼 종실은 수발아시 a-amylase 등의 활성 증가로 종실의 주 구성성분은 전분을 분해하여 전분의 치밀한 결정구조가 와해된 분상질립이 발생한다(Zhu et al., 2019). 이 분상질립은 아밀로스함량이 낮고 유리당함량이 높은 특성을 지니고 있는데, 이는 정상종자 대비 전분호화특성의 감소를 불러온다(Lin et al., 2020). 수발아 발생정도에 따라서 최고, 최종 최저 점도가 감소해 호화특성이 감소하였지만, 치반점도는 감소해 노화속도가 낮아지는 것처럼 보이는 결과를 나타내었다(Fig. 6). 하지만 이를 통해 호화특성이 감소하였지만 노화에 대한 지표가 좋아졌다고 하기 어려우며 이러한 결과는 추가적인 실험을 통해 구명할 필요가 있을 것으로 생각된다. 등숙기 온도 증가에 따라서 치반점도가 감소했지만 노화특성이 좋아지는 것이 아니었다는 다음과 같은 기존의 결과가 있다. 전분호화특성의 결과에 큰 영향을 미치는 아밀로스 및 아밀로펙틴의 함량, Chain-length 등의 특성은 등숙기 온도에 따라서 크게 좌우되는데, 등숙기 온도가 증가함에 따라서 아밀로스 함량이 감소하게 되면, 종실에 열과 수분을 가하였을 때 호화되어 부풀어 오르는 특성과 관련된 최고, 최종점도가 증가하고, 노화특성과 양의 상관관계를 나타내는 치반점도는 감소한다(Tester & Morrison, 1990; Chun, 2009). 하지만 노화특성과 양의 상관관계인 길이가 상대적으로 긴 아멜로펙틴 Chain 등의 비율이 고온등숙이 되는 경우 증가하게 되는데, 이에 따라서 고온등숙이 되어도 노화특성에 영향을 미치는 두 요인인 아밀로스 함량과 아멜로펙틴 Chain이 서로 상반된 영향을 주게 되어 치반점도가 낮아졌음에도 결국 노화정도는 감소되지는 않는다고 한다(Vandeputte et al., 2003; Chun, 2009). 한편 단백질의 경우에는 기존의 일부 보고에서는 수발아에 따른 단백질의 함량 변이가 없었다고 보고하였는데(Kim et al., 2008; Yao et al., 2020), 이와는 다르게 본 시험의 결과에서는 일정 수발아율 이상부터는 감소하는 결과를 보였으며(Fig. 7), 이는 발아 시 침지 시간이 일정 시점이 지남에 따라 단백질 함량이 감소하였다는 기존의 보고된 결과와 같았다(Palmiano & Juliano, 1974). 이는 수발아 발생 초기에는 발아에 사용하기 위한 에너지원으로 1차적으로 유리당 및 전분 등 탄수화물의 이용 정도가 상대적으로 많고(Palmiano & Juliano, 1974) 단백질 함량 감소 정도가 작아 비율상 단백질함량의 감소가 없었던 것처럼 결과가 나타난 것으로 생각된다. 이렇듯 수발아가 발생된 종실을 쌀로 취반하여 섭취할 경우에는 식미와 관련된 품질특성이 감소하게 된다. 하지만 발아현미의 경우 종자가 발아 시 GABA Gamma-Oryzano, 스쿠알렌 등 기능성 물질이 생성되는 것을 이용하여, 이 물질들을 추출하여 기능성식품을 개발하기도 한다(Cho et al., 2011). 추후 이러한 수발아가 발생된 종실의 유용기능성 물질을 분석하고 이를 활용 할 수 있는 방법을 고안하여, 농가에서 수발아 발생 종자를 활용 할 수 있는 방법을 제시하는 추가적인 실험이 검토될 필요가 있을 것으로 생각된다.
적 요
등숙기간 동안의 온도에 따라서 동일한 출수 후 일수가 경과하더라도 출수 후 적산온도가 달라져 종실의 등숙단계가 달라지기 때문에 수발아에 영향을 줄 수 있는 종실의 특성이 달라지게 된다. 그렇기 때문에 수발아 검정을 위한 처리 시점은 출수 후 일수가 아닌 출수 후 특정 적산온도에 조건을 맞춰 시험을 진행해야 한다. 한편 수발아 발생은 등숙기간 동안의 온도에 따라서 크게 달라진다. 이에 따라서 발생하는 전분립의 구조, 전분함량 및 ABA함량의 변이 등은 수발아 발생에 영향을 줄 것으로 생각되며, 특히 고온 등숙 시 수발아 발생율의 증가와 종실 전분립의 구조, 유리당 함량 등 종실 이화학적 특성 변이와의 관계에 대한 추가적인 검토가 필요할 것으로 생각된다. 또한 고온등숙시 수발아 발생율이 증가하기 때문에 등숙기 고온에 노출되지 않도록 이앙시기를 조절하여 수발아 발생위험을 감소시켜야 한다.
