

Introduction
MATERIALS AND METHODS
Experimental materials and treatment conditions
Analytical and Statistical Methods
RESULTS
Temperature-dependent variation in heading date by growth stage
Temperature-dependent variation in growth rate by growth stage
Major temperature parameters influencing the heading date by growth stage
DISCUSSION
Effect of temperature on heading during the photo- sensitive period
Discrepancy between heading and elongation response to temperature
Daily temperature difference affect heading response during photo-sensitive period
CONCLUSIONS
Introduction
Global warming and frequent extreme weather events are likely to pose serious threats to stable rice production and supply by increasing the risk of serious adverse impacts on plant growth and development (Kotak et al., 2007). In particular, variations in the heading date of rice ascribable to extreme weather events including abrupt shifts in average temperature, especially heatwaves, can cause seed infertility in the heading-flowering stage (Satake & Yoshida, 1978; Wang et al., 2019). Similarly, changes in weather conditions during the ripening stage can have a serious impact on the yield of rice and quality of harvest (Shi et al., 2016). Therefore, investigating the effect of weather conditions on heading response will provide important basic data for developing technologies or new cultivars capable of stabilizing rice quality and yield by resisting or adapting to the changing climate. The two major environmental factors determining the heading response of rice are photoperiod and temperature (Ahn & Vergara, 1969; Dua et al., 1990; Collinson et al., 1992). Rice copes with short photoperiods by activating the expression of florigen genes which induce panicle initiation by moving to the shoot apical meristem in leaves (Bernier & Perilleux, 2005; Corbesier et al., 2007; Tamaki et al., 2007). To shed light on the control mechanism of such responses responsible for the developmental transition from vegetative to reproductive growth, extensive research has been undertaken with various foci including photoperiod-dependent heading response, photosensitivity- related gene expression mechanism, and quantitative trait locus (QTL) analysis (Yano et al., 1997; Yamamoto et al., 2000; Nishida et al., 2001; Yang et al., 2018). However, the effects of temperature on heading response have not been sufficiently studied to date (Luan et al., 2009; Kovi et al., 2013). Studies on temperature-dependent heading response have been conducted at the molecular level, investigating the effect of temperature on promoting or inhibiting panicle initiation control mechanism to cope with short photoperiod during a specific period, without however, establishing a direct link to temperature because plants were at different growth stages during the sampling period (Lee et al., 2011; Luan et al., 2009; Song & Luan, 2012; Song et al., 2012; Song et al., 2013). Although the effect of temperature on the heading response control mechanism has been separately studied, no results capable of elucidating the interconnected control mechanism have been reported to date (Kovi et al., 2013). On the contrary, a number of studies have been conducted to analyze temperature-dependent heading response under a fixed photoperiod (Vergara & Chang, 1985; Roberts & Summerfield, 1987), and it has been reported that the pre-heading growth period decreases nonlinearly as the temperature increases within a certain temperature range (Yoshida, 1981; Haniu et al., 1983; Loomis & Connor, 1992). It was found that higher the minimum temperature for the same average temperature i.e., smaller the daily temperature difference, lower the growth rate (Yin et al., 1996), and that the effect of temperature on heading varies depending on the stage of growth (Wang, 1960; Yin et al., 1997). However, earlier experiments on the effect of temperature on different growth stages yielded totally different interpretations (Ahn & Vergara, 1969; Shibata et al., 1969; Yin et al., 1997). In this study we aimed to shed light on the effect of temperature change on heading response depending on the growth stage and ecotype of rice grown in the natural photoperiod conditions with naturally changing length of photoperiod. To this end, we investigated the association between plant growth pattern and heading variation under different treatment conditions and analyzed the effects of major temperature parameters, such as minimum, average, and maximum temperature, and daily temperature difference, on heading response in each treatment period using random forest analysis.
MATERIALS AND METHODS
Experimental materials and treatment conditions
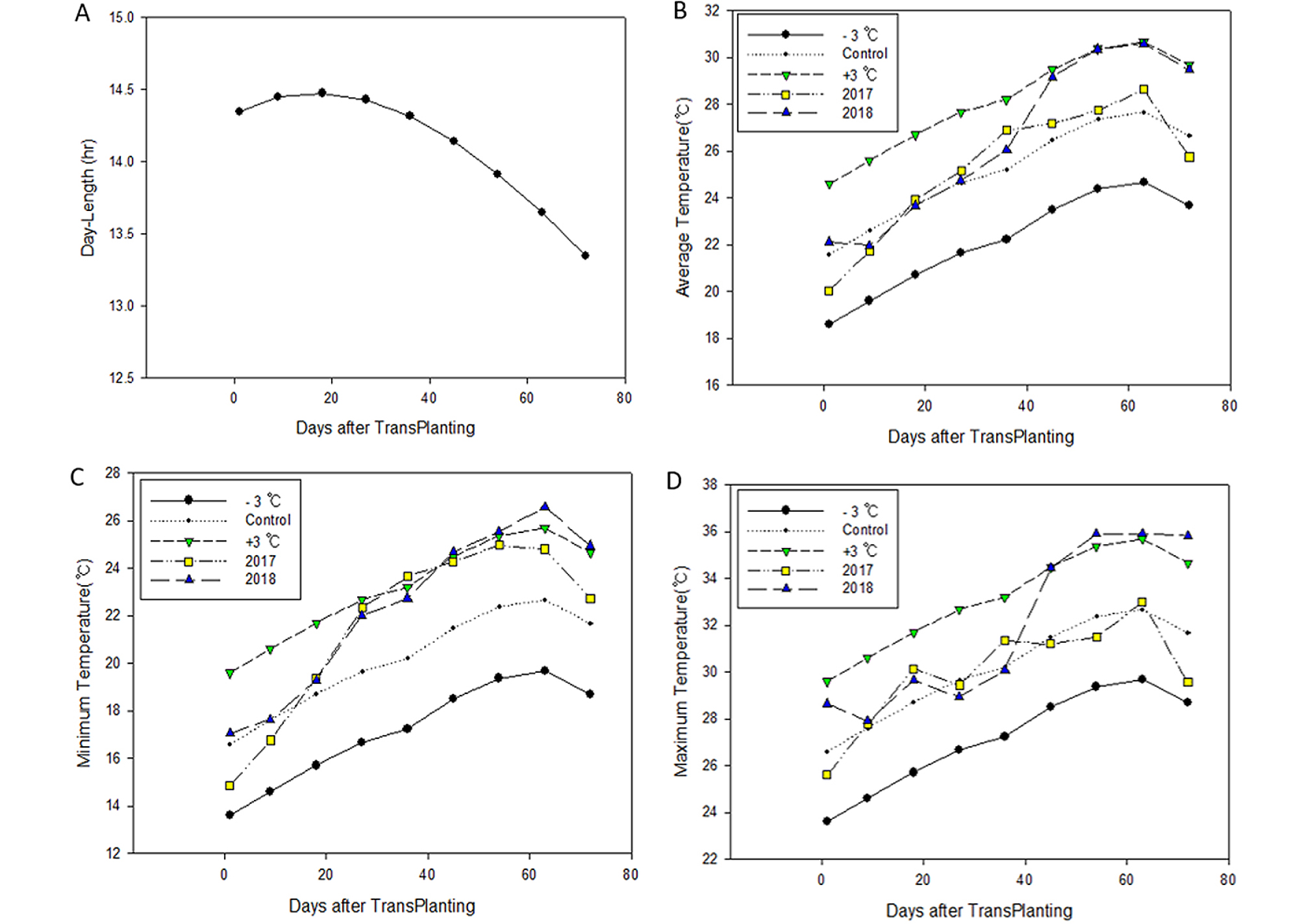
For our experiment, we used six rice (Oryza sativa) cultivars (temperate japonica type), two each for early maturing (Odae & Ungwang), medium maturing (Gopum & Daebo), and mid-late maturing (Sanuri & Shindongjin) ecotypes. For all cultivars, 20-day-old seedlings were transplanted on the same day (May 30) in 2017 and 2018 using 1/5000а Wagner pots, with three plants per pot. As a fertilizer, we used a composite slow-release fertilizer based on 9 kg/10a nitrogen, 4.5 kg/10a phosphate, and 5.7 kg/10a potassium at the area ratio corresponding to three plants (planting distance: 30 × 14 cm) instead of the pot area. Seedlings were sown and grown in 406-cell seedling trays. The average number of leaves on the transplantation day (May 30) was 3.9. Temperature treatment was applied to three groups of freshly transplanted rice plants in thermo-regulated glass rooms of the artificial weather facility of the National Institute of Crop Science: the control group was treated with temperatures (hourly automatic temperature control) varying at 9-day intervals in each treatment period based on the 15-year (2001–2016) average temperature in Jeonju area; the low- and high-temperature groups were treated with temperatures 3°C lower or higher than those applied to the control group (Fig. 1). Except for the temperature treatment period of nine days, the rice plants were grown in natural weather conditions.
Analytical and Statistical Methods
For the heading date analysis, “heading date” was defined as the number of days from transplanting to the appearance of the first spikelet, with nine replicates for each treatment group, as counted between 13:00 h and 14:00 h on a daily basis. The importance index of temperature on heading response in each growth stage was calculated through the following steps: (1) the temperature-dependent heading date variation at 9-day intervals was checked, (2) a value of +1 or –1 was assigned to one-day delay or advance, respectively, compared with the control group, (3) the heading date variation in response to a difference of 1°C in the daily average temperature during each treatment period was summed, (4) the total value was divided by the extent of variation in the corresponding period, (5) the value was expressed as a percentage. The vegetative growth phase was divided, based on the growth response of the control group grown in natural weather conditions, into the rooting stage (from transplanting to the emergence of the first leaf; ~9 days) and the early tillering stage (from the primary tillering stage, with the emergence of the primary tiller, to the secondary tillering stage with the emergence of the secondary tiller from the first node position; 7.5 leaves, ~9 days). The reproductive growth phase is generally considered to be completed in 35 days but varies depending on the criteria for determining the heading date (Vergara et al., 1965; Ikeda, 1975). In our experiment, given that the heading date was determined as the time when 50% of the panicles became exserted from the boot, we calculated panicle initiation, spikelet differentiation, and meiosis stages to be 30, 24, and 15 days prior to heading, respectively (Matsubayashi, 1963; Katayama, 1971).
The temperature-dependent variation in plant height prior to heading was computed using the following logistic model (Black & Leff, 1983):
| $$E=E_{min}+\frac{E_{min}+E_{max}}{1+(t/t_h)^{-Er}}$$ |
where E is plant height (cm), t is the number of days up to heading, Er is the curve pattern, which determines the rate of elongation reduction, Emax and Emin are the maximum and minimum elongation rate, respectively, and th is the time when 50% of Emax is reached. Er, Emax, Emin, andth are coefficients determined by nonlinear regression analysis, which was performed using SigmaPlot 11.1.
Results of each experiment were analyzed using the R Statistical Package (Version 3.2.2), with the differences considered to have statistical significance at p < 0.05. Random forest analysis was performed using the R-package randomForest and caret. The values of mtry and ntree, which determine the number of branches for explanatory variables and each node of the decision tree model, were set to those yielding the lowest OOB errors as follows, using tune RF and gridsearch: for the early maturing cultivars, mtry = 4, OOB = 0.93, and ntree = 30000; for medium maturing cultivars, mtry = 4, OOB = 2.15, and ntree = 10000; for the mid-late maturing cultivars, mtry = 5, OOB = 3.08, and ntree = 10000.
RESULTS
Temperature-dependent variation in heading date by growth stage
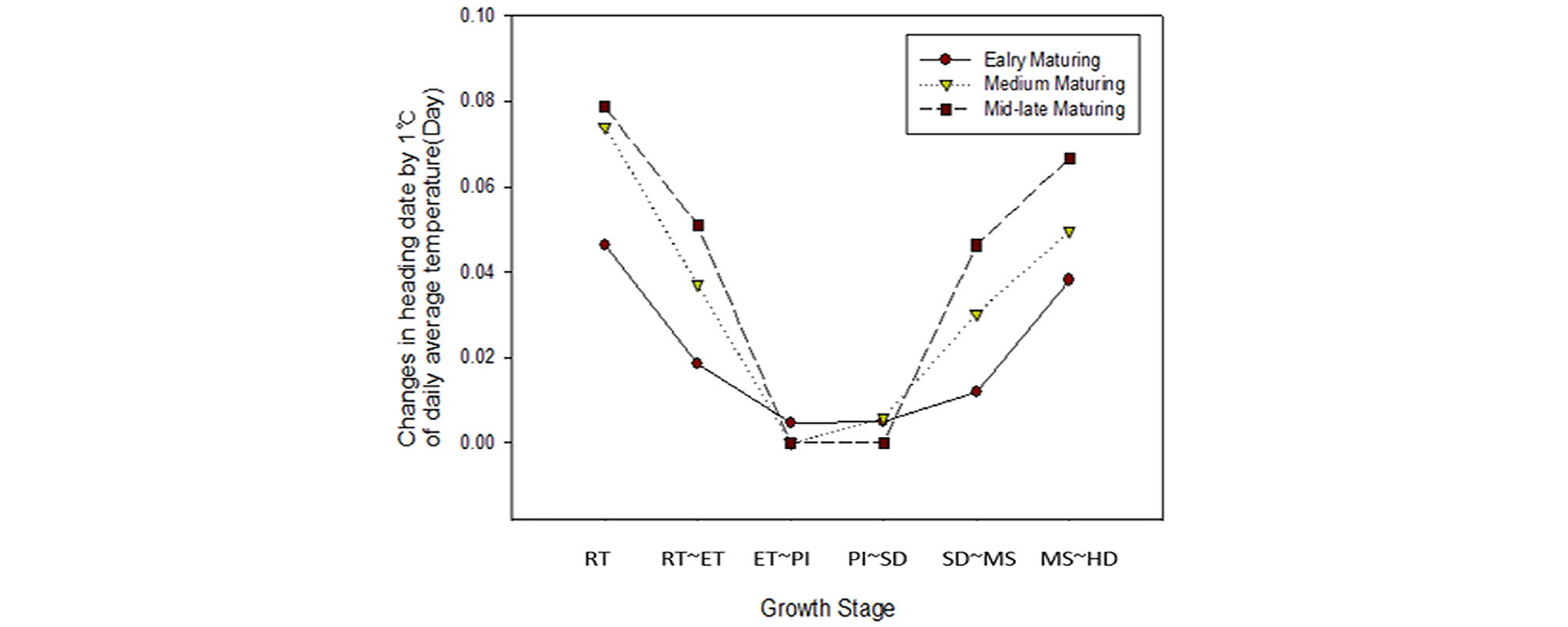
As shown in Fig. 2, the influence of temperature on heading date varied by growth stage. In all ecological types, increase in temperature nonlinearly shortened the heading date in some growing stages, but there were also stages where no significant changes were observed, especially in the period between 36 and 27, 45 and 27, and 51 and 24 days before heading for early maturing, medium maturing, and mid-late maturing cultivars, respectively. Table 1 presents the magnitude of temperature effect on the heading date in different treatment periods as the difference in the days to heading induced by 1°C increase in the daily average temperature. The magnitude of temperature-dependent variation in heading date increased with an increase in the length of growth period, i.e., in the order of early maturing, medium maturing, and mid-late maturing ecotypes. The same trend was observed in the growth stages nonresponsive to temperature change. The growth period of rice plants is divided according to the days before heading: rooting, early tillering, panicle initiation, spikelet differentiation, and meiosis stages (Matsubayashi, 1963; Katayama, 1971). Fig. 3 illustrates the importance index in each growth stage. In all cultivars and ecotypes, the highest importance index was demonstrated by the rooting stage, followed by meiosis, early tillering, spikelet differentiation, and panicle initiation stages. In particular, during the photo-sensitive period (PSP), namely around the panicle formation time, the effect of temperature change on heading response was greatly reduced, which supports the finding of Ahn and Vergara (1969) that the effect of temperature was insignificant during PSP.
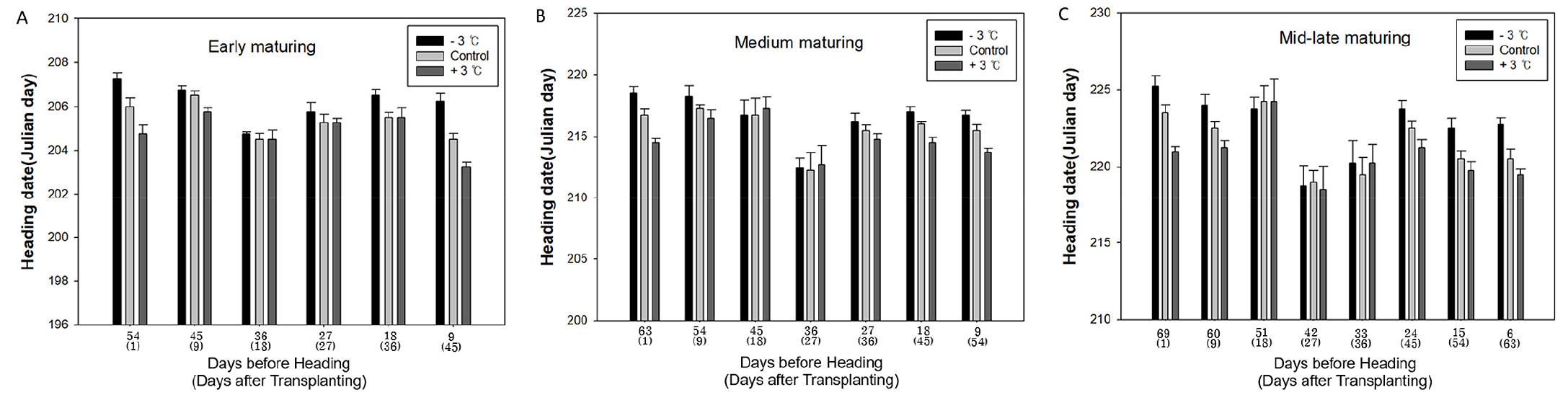
Table 1. Effect of degree of temperature on heading by days before heading.
Temperature-dependent variation in growth rate by growth stage
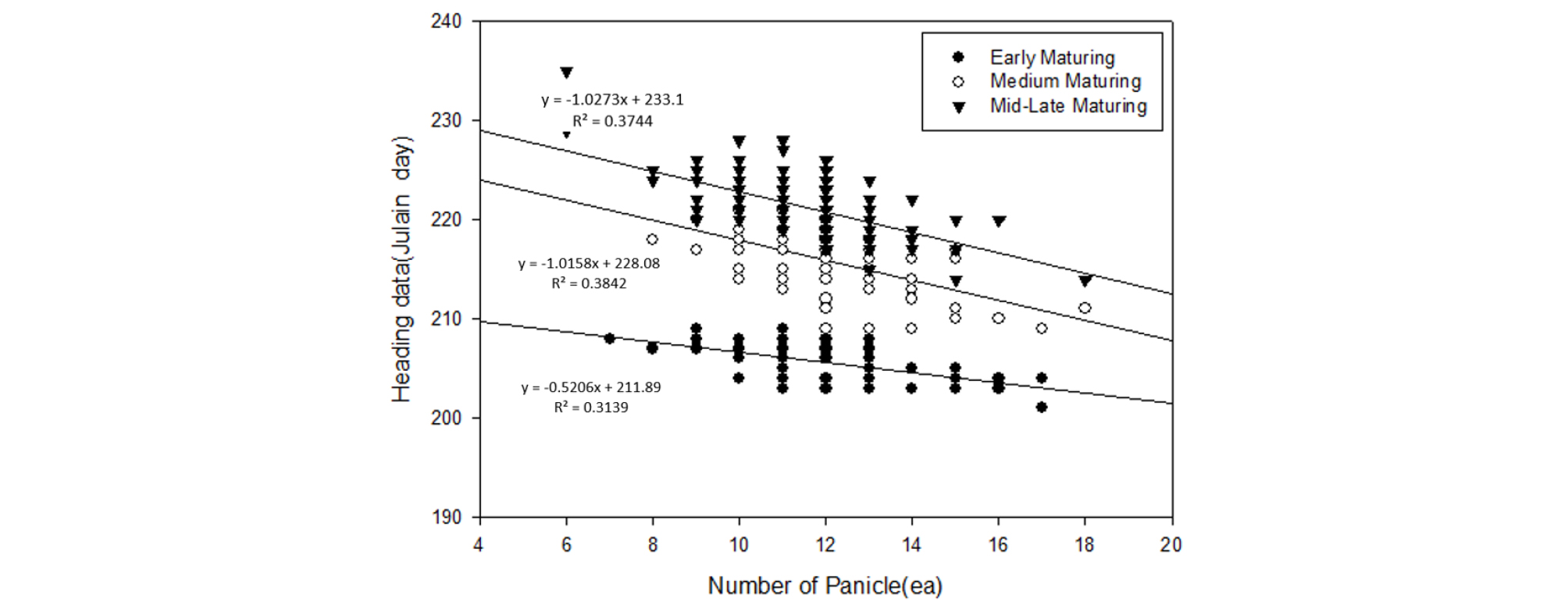
Fig. 4 shows the temperature-dependent variation in plant height growth rate by growth stage. No significant inter- ecotype differences were observed in the plant height growth rate after the 9-day temperature treatment in each growth stage. The average values (± SD) of plant height growth during the entire growth period in the low, normal, and high temperature conditions were 6.5 (± 1.2), 12.6 (± 2.1), and 17.3 (± 2.3) cm, respectively. This difference in growth response was maintained at similar levels until a specific period and sharply decreased after the peak growth period (Th in Table 2), which appeared earlier in the high- temperature treatment group. Contrary to the decrease in the effect of temperature on heading response around PSP, the effect of temperature on growth rate was maintained during PSP and began to decrease after the spikelet differentiation stage. Whereas, the correlation analysis between heading date and post-heading growth components, such as clum length, panicle length, number of spikelets per panicle, and number of panicles per plant, revealed a negative correlation only between the number of panicles and heading date (Fig. 5).
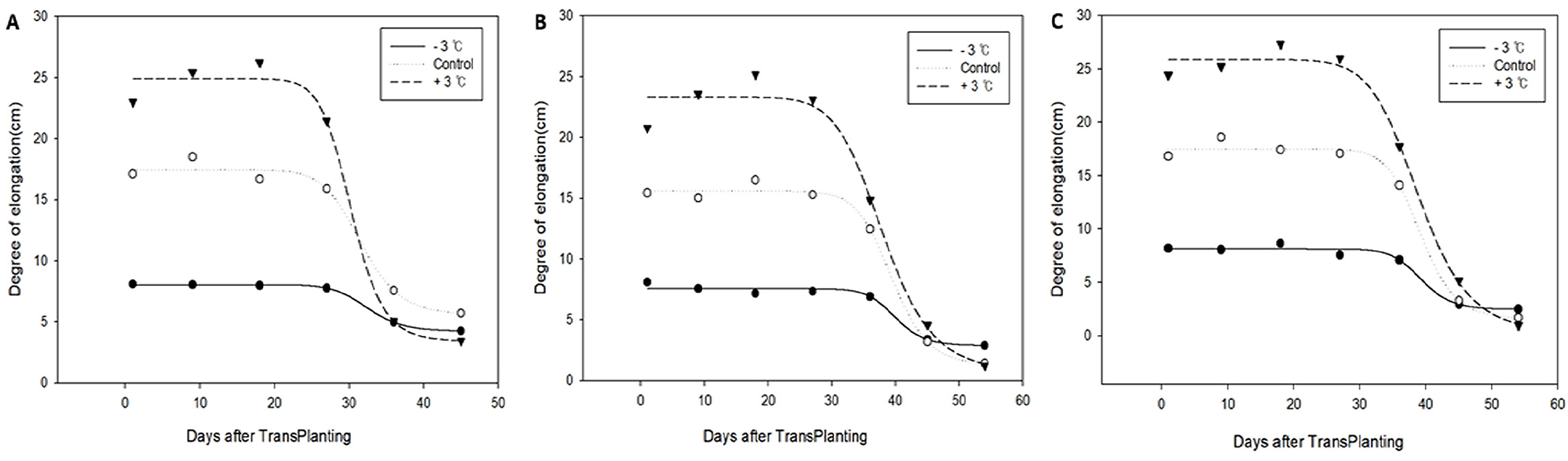
Table 2. Parameters of the logistic function used to describe elongation response in relation to the days after transplanting under three temperature regimes during the growth period from transplanting to heading.
- "*" and "**" represent significance at the 0.05 and 0.01 probability levels, respectively.
Major temperature parameters influencing the heading date by growth stage
Table 3 presents the results of the correlation analysis between heading date and temperature parameters (minimum, average, and maximum temperatures and daily temperature difference) by treatment group during the growth period at 9-day intervals. In the table, the five highest correlation coefficients between temperature parameters and growth stage are listed. These correlation analysis results showed trends similar to that shown by the importance indices by growth stage (Table 1). Given that various environmental factors have complex interacting effects on the heading response, multiple regression analysis needs to be performed for proper interpretation. However, multiple regression analysis inevitably leads to collinearity problem among minimum, maximum, and average temperatures in each treatment period, making it impossible to establish a formula using all factors as variables. In contrast, random forest analysis is better suited for highly interacting complex data using many explanatory variables with variable importance (Breiman, 2001). Fig. 6 shows the importance indices of the aforementioned temperature parameters influencing heading date resulting from random forest analysis. In the case of the early maturing cultivars, the five highest importance indices were demonstrated by the temperature parameters in the rooting and meiosis stages (1st and 6th treatment periods), similar to the results of the correlation analysis. In the cases of medium and mid-late cultivars, however, the daily temperature difference in PSP (3rd and 4th treatment periods), which is characterized by a reduced effect of temperature on heading response, was included in the group of five highest importance indices along with the temperature parameters in the 1st treatment period. This result is consistent with that of the experiment conducted by Shibata et al. (1969) according to which the daytime temperature 29 and 32 days before heading had an insignificant effect on heading, but the nighttime temperature greatly influenced it. Figs. 7, 8 present the five major factors affecting heading date in terms of importance index in early maturing. While higher temperature tended to accelerate heading date in the early maturing ecotype, the medium maturing ecotype did not show a negative correlation between the 4th-period average temperature and heading date, and higher daily temperature difference accelerated heading date. In the medium maturing ecotype as well, higher daily temperature difference was associated with an earlier heading date with the increase in the daily temperature difference during PSP (4th treatment periods).
Table 3. Correlation coefficient between heading date and weather factors by temperature treatment at different growth stages.
- MNT: Minimum temperature, AVT: Average temperature, MXT: Maximum temperature, DTD: Daily temperature difference, WP: Whole growth period. DBH: Days before heading. DAT: Days after transplanting
- Information on the relationship between temperature treatment periods and growth stages can be found in Materials and Methods, Table 1.
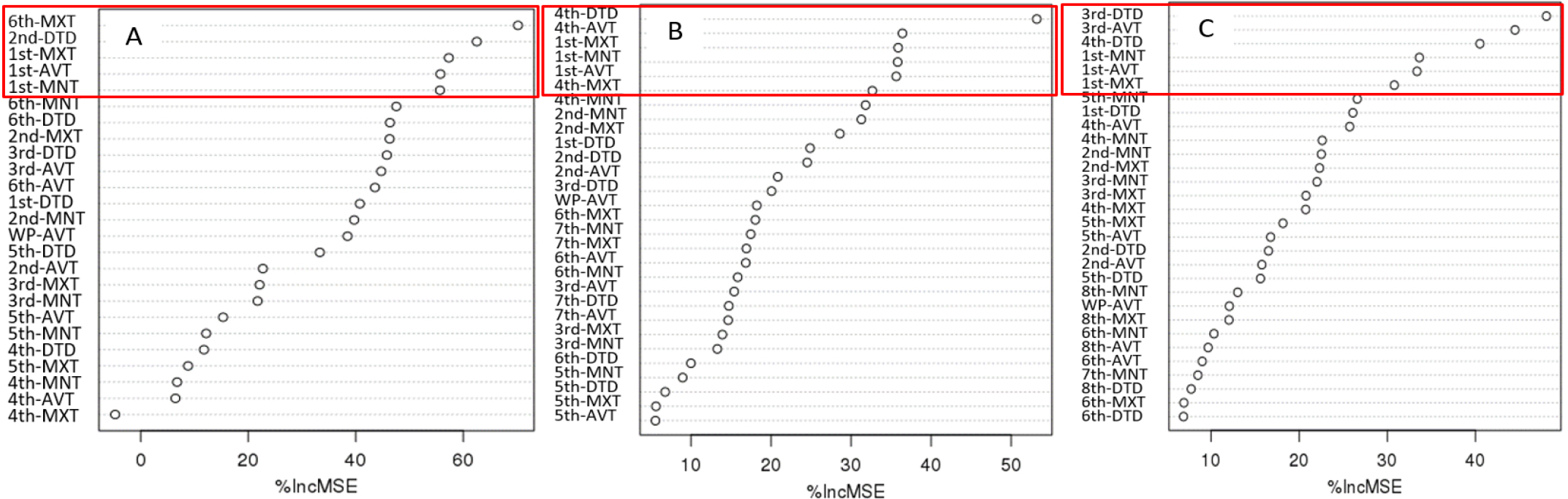
Fig. 6.
Importance index (% increase MSE) of weather factors influencing heading through random forest analysis. A: Early maturing type. B: Medium maturing type. C: Mid-late maturing type. MNT: Minimum temperature, AVT: Average temperature, MXT: Maximum temperature, DTD: Daily temperature difference, WP: Whole growth period. Information on the relationship between temperature treatment periods and growth stages can be found in Materials and Methods, Table 1.
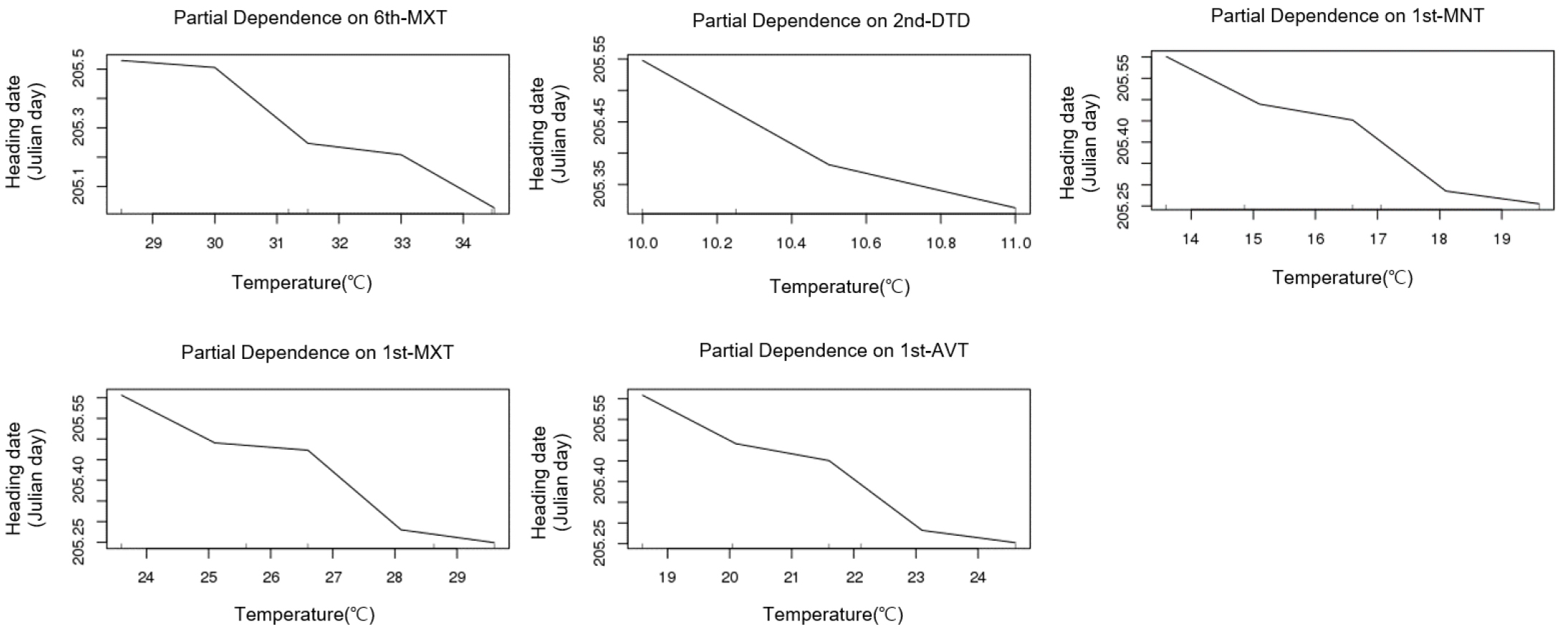
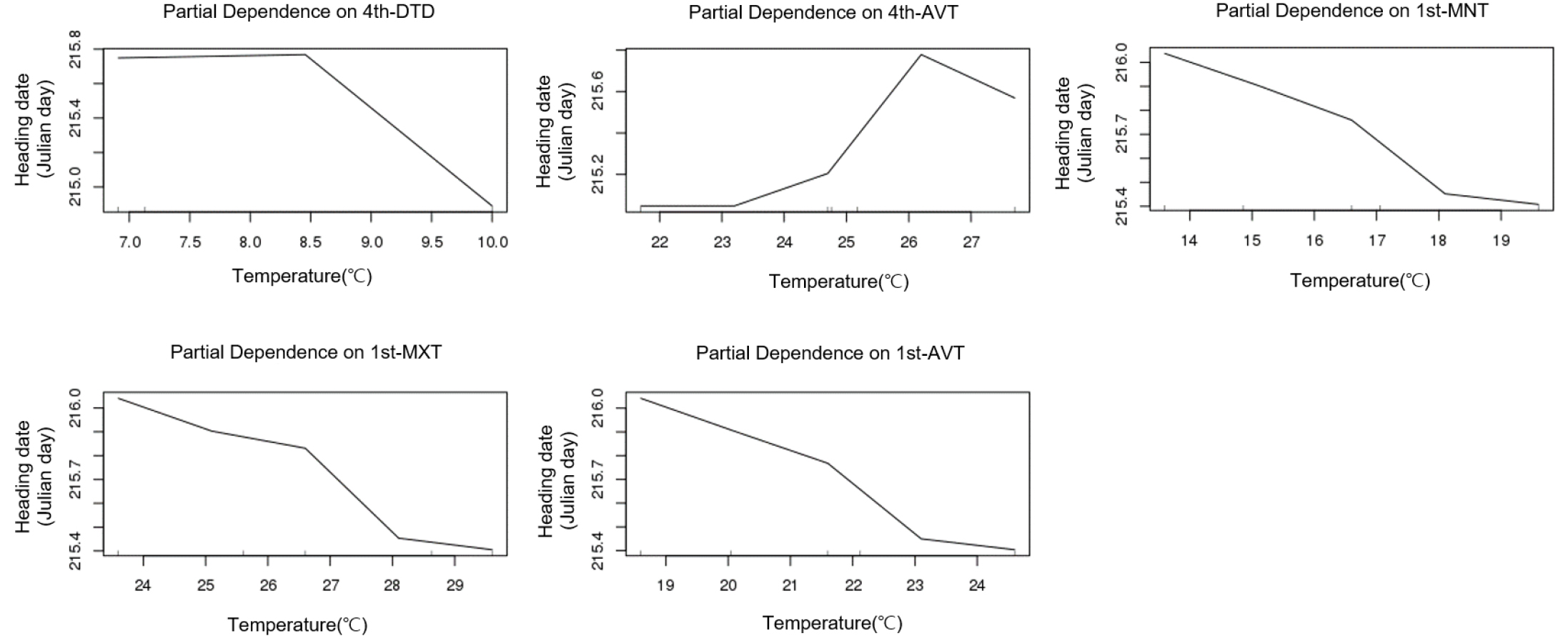
DISCUSSION
Effect of temperature on heading during the photo- sensitive period
Photoperiod and temperature are two major environmental factors influencing the heading response of rice (Ahn & Vergara, 1969; Dua et al., 1990; Collinson et al., 1992). Vergara et al. (1985) & Roberts et al. (1987) divided the pre-heading period (from germination to heading) into photo- insensitive, photo-sensitive, and again photo-insensitive stages. In the experiment by Yin et al. (1997), these were renamed as basic vegetative period (BVP), photo sensitive period (PSP), and post photo-sensitive period (PPP), respectively, for convenience. According to previous findings regarding temperature and photoperiod, photoperiod influences heading only in PSP (Mimoto et al., 1989; Collinson et al., 1992), and temperature throughout the entire growth period (Yoshida, 1981), but to a varying extent depending on the growth stage (Wang, 1960; Yin et al., 1997). However, results of the experiments conducted to date regarding the varying effects of temperature on heading between BVP and PSP yielded completely different interpretations. While Yin et al. (1997) reported that heading response is more sensitive to temperature in PSP than in BVP and PPP, Ahn and Vergara (1969) reported that temperature has a great effect in BVP and a negligible effect in PSP. Shibata et al. (1969) reported that heading response was sensitive to daytime temperature 11 days before heading, but neutral to daytime temperature 29 and 32 days before heading. Similar to the latter two results, the results of our experiment revealed that the effect of temperature was high during the early growth stage after transplanting, negligible up to the spikelet differentiation stage, which is considered a PSP, and increased again up to the meiosis stage till heading (Fig. 3, Table 1). The contradictory conclusions derived by the researchers from their experimental results regarding the effect of PSP temperature are presumably due to different photoperiod conditions. While Yin et al. (1997) investigated the response to temperature in an experimental setup of fixed short- photoperiod conditions (12-h light/12-h dark), the experiments by Ahn & Vergara (1969) & Shibata et al. (1969) were conducted in the natural photoperiod conditions (14-h 30-m light/9-h 30-m dark), similar to those in our experiment. This can also be inferred from the results of the experiment conducted by Yoshida & Hanyu (1964) that the photoperiod critical to heading response is dependent upon temperature. Furthermore, Komiya et al. (2009) reported that panicle initiation is caused by RFT1 (Rice Flowering Locus T 1) in long-photoperiod conditions and by RFT1 and Hd3a (Heading date 3a) in short-photoperiod conditions. RFT1 is the closest homolog of 13 FT-like genes in rice to Hd3a. & Komiya et al. (2008) reported that RFT1 promotes flowering in the absence of Hd3a. and it act as an integrator of multiple signals under long day conditions with Ehd1. This suggests that different heading response mechanisms are activated depending on photoperiod conditions, and further studies are necessary to compare the effect of temperature on heading response in long- and short-photoperiod conditions. The conclusion drawn from the experimental results by Summerfield et al. (1992), according to which temperature and photoperiod do not interact in their impact on heading, may be ascribable to their experimental treatment conditions, which were inappropriate to draw a conclusion applicable to photoperiodic variation, because they did not compare the temperature treatment conditions in different photoperiod conditions.
Discrepancy between heading and elongation response to temperature
Unlike heading response, degree of elongation response increased with the increase in temperature until before the spikelet differentiation stage regardless of the growth stage, and no temperature-dependent difference in growth rate was observed afterwards (Fig. 4). In particular, in contrast to heading response, elongation response sensitivity to temperature was observed in PSP treatment. These results allow the assumption that vegetative growth rate can be predicted in temperature conditions within a specific range but predicting heading response based on them may be prone to error. From the finding that individuals with a higher number of panicles per plant showed an accelerated heading date (Fig. 5), it can be inferred that individuals that grow faster until the photoperiod-induced sensitivity to short daylength produce a larger number of tillers, resulting in faster heading. This can also be inferred from the report of Tsuji et al. (2015) that Hd3a, which regulates heading response, promotes tillering.
Daily temperature difference affect heading response during photo-sensitive period
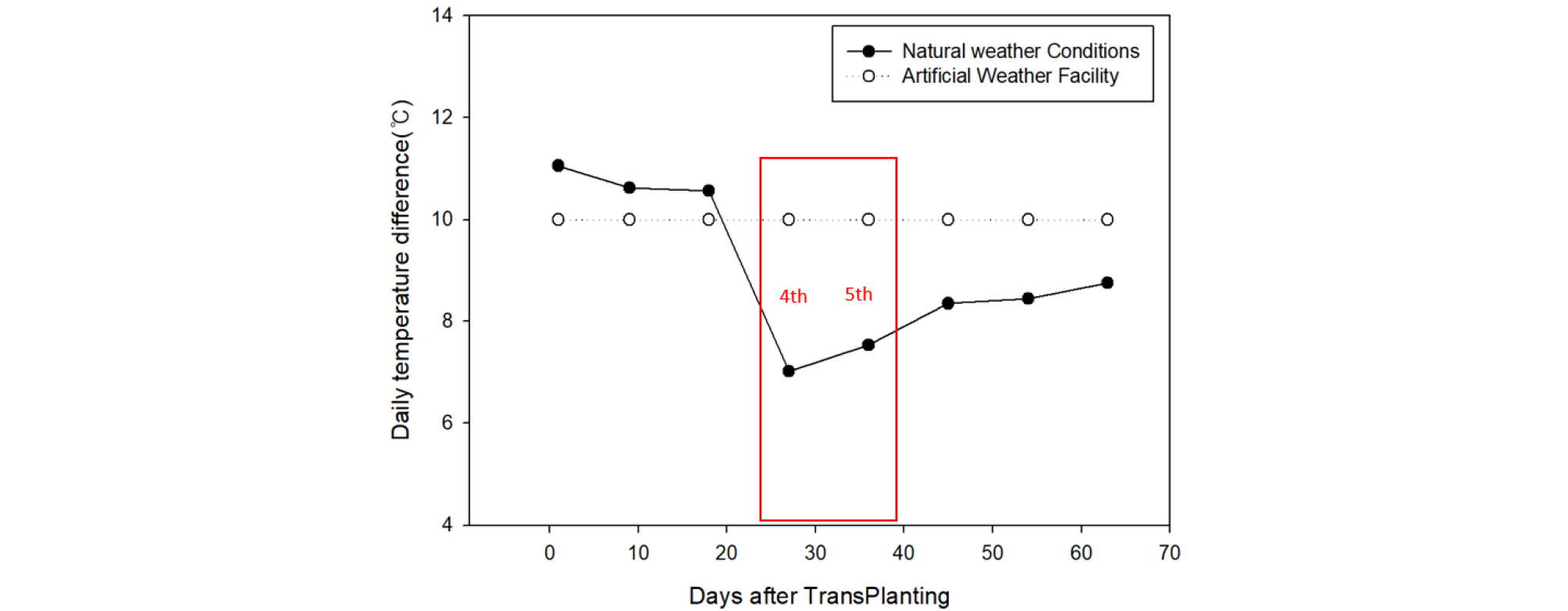
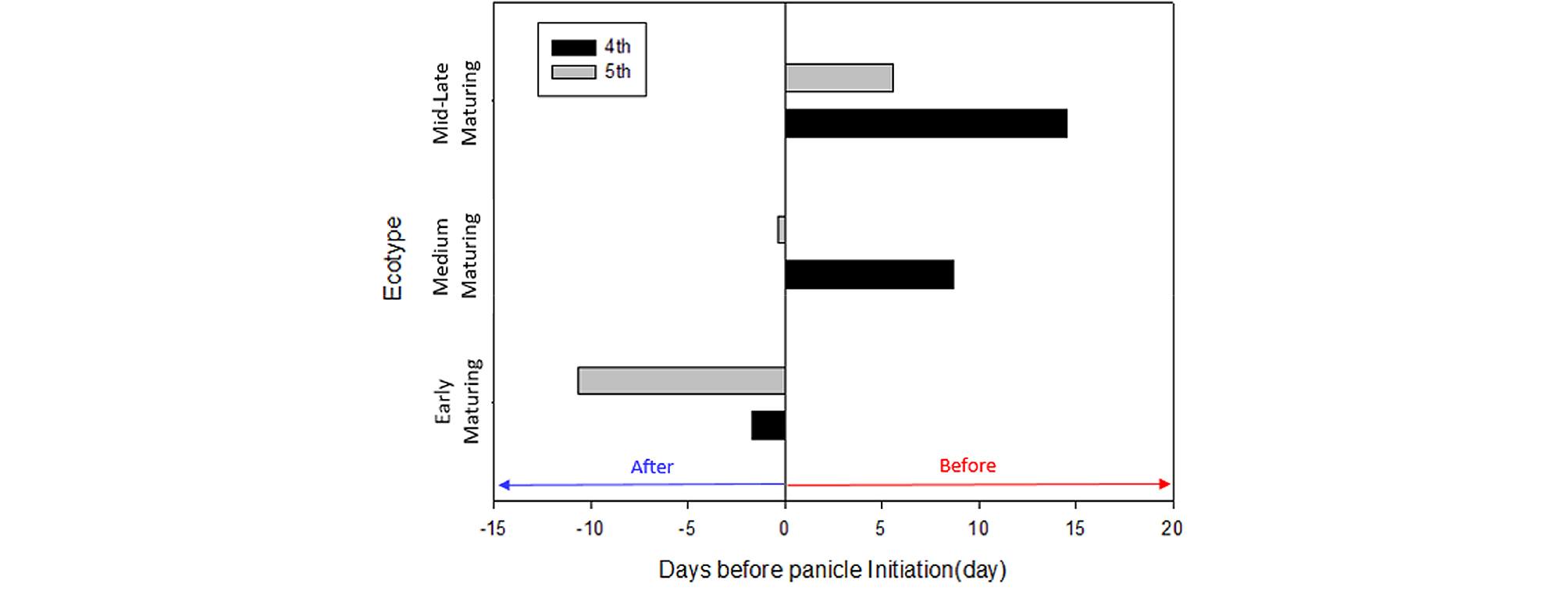
A comparison of the average heading date between treatment periods revealed that while temperature treatment does not induce any significant differences in heading response, there are periods associated with a faster average heading date, namely the 4th treatment period in the medium maturing ecotype and the 4th and 5th treatment periods in the mid-late maturing ecotype (Fig. 2). In the results of random forest analysis as well, the importance index of daily temperature difference in the 4th treatment was the highest. This may be explained by the sudden decrease in daily temperature difference in the natural weather conditions due to the rise in minimum temperature in the 4th treatment period shortly after the summer solstice, whereas the daily temperature difference was maintained at 10°C in the artificial weather facility (Fig. 9). Comparing the growth stages among the maturing ecotypes, both the 4th and 5th treatment periods fell after panicle initiation in the early maturing cultivars, ~9 days before (in PSP) and after panicle initiation, respectively, in the medium maturing cultivars, and ~15 and ~6 days before panicle initiation (in PSP), respectively, in the mid-late maturing cultivars (Fig. 10). From these results, it can be inferred that daily average temperature has little impact on heading response during PSP prior to panicle initiation in normal growth conditions, but the nighttime minimum temperature and daily temperature difference affect heading response during PSP. Growth response to daily temperature difference over the entire growth period was demonstrated in the experiment conducted by Yin et al. (1997). Also, considering the experimental finding by Shibata et al. (1969) that the daytime temperature 29 and 32 days before heading had an insignificant impact on heading, but the nighttime temperature greatly influenced it, there is a need to carry out an in-depth study on the effect of daily temperature difference on heading response in PSP.
CONCLUSIONS
The following gives an overview of the key results regarding the temperature-dependent heading response of rice by growth stage and related explanations and interpretations:
1. The effect of temperature on heading response varies depending on the growth stage. It is the highest in the rooting stage, followed by meiosis, early tillering, spikelet differentiation, and panicle initiation stages; in particular, the effect of temperature in the photo-sensitive period (PSP) was verified to be negligible.
2. The analysis results for the temperature parameters influencing heading by growth stage revealed the change in daily temperature difference in PSP as one of the main factors, in addition to the temperature in the rooting and meiosis stages, and an in-depth study of this aspect is considered necessary.
3. Comparing the temperature-dependent growth response by growth stage, the temperature-dependent difference in growth response was maintained up to the panicle initiation period, but temperature treatment had little effect on growth response afterwards, which did not coincide with heading response.
