

서 론
재료 및 방법
공시재료 및 콩 발아
발아 중 물리적 요인 처리
이소플라본 함량 분석
통계분석
결과 및 고찰
물리적 요인이 발아 중 이소플라본 함량에 미치는 영향
콩 발아와 이소플라본 종류별 함량 변화
적 요
서 론
이소플라본은 주로 두과식물에 존재하는 이차대사물질로서 특히 콩에 상당량이 함유되어 있다. 이소플라본은 항산화 및 항암 효과 등 여러 성인병에 대해 예방적 혹은 치료적 효과가 있는 것으로 알려져 최근까지도 이와 관련된 많은 연구가 이루어지고 있다(Nielsen & Williamson, 2007; Messina, 2014; Dhayakaran et al., 2015; Ko et al., 2017). 그러나 이소플라본 함량은 유전적(Wang & Murpy, 1994; Gutierrez-Gonzalez et al., 2010) 및 환경적 요인(Kim et al., 1996; Gutierrez-Gonzalez et al., 2009)에 의해 영향을 많아 받는 까닭에 식물체내 이소플라본의 합성 및 축적에 관한 연구는 앞으로도 계속 수행되어야 할 과제이다.
콩의 이소플라본 함량에 영향을 끼치는 환경적 요인으로는 등숙기의 온도(Lee et al., 2008) 및 수분공급(Bennett et al., 2004; Gutierrez-Gonzalez et al., 2010)등에 관한 보고가 있다. 또한 최근에는 이소플라본 축적과 관련된 유전자가 속속 밝혀지고 있으며(Yoneyama et al., 2016; Yuk et al., 2016; Ahmad et al., 2017) 이에 관련된 연구도 많이 수행되고 있다. Gutierrez-Gonzalez et al.(2010)은 식물체내 이소플라본 축적에는 isoflavone synthase genes (IFS1 and IFS2)가 많은 영향을 미치며 특히 스트레스 조건 하에서는 IFS2의 발현이 중요한 역할을 한다고 하였다. Yu et al. (2000)은 비 콩과식물에 IFS 유전자를 형질전환시킴으로써 이소플라본이 축적됨을 확인하는 한편 형질전환체 유묘에 UV를 조사하니 이소플라본 함량이 높아진다고 하였으며, Li et al. (2016)은 형질전환체를 사용하여 이소플라본 생합성과정에 관여하는 유전자를 탐색한 바 있다. 이밖에도 이소플라본과 관련된 QTLs을 분석한 결과 이소플라본 축적에는 2개 이상의 loci에서 epistasis 등 유전자 간 상호작용 뿐만 아니라, 유전자의 minor effects가 존재하는 등 수 많은 요인이 관여된다고 하였다(Gutierrez-Gonzalez et al., 2009).
콩에서 이소플라본은 뿌리, 유묘, 꽃 등에 존재하지만 특히 종자와 잎에 풍부하게 함유되어 있다(Gutierrez-Gonzalez et al., 2010). 따라서 콩의 발아를 통한 이소플라본 축적 등에 관한 연구도 수행되고 있다(Yun et al., 2013; Yang et al., 2015; Lee et al., 2018). 한편, Yuk et al. (2016)은 R3 시기의 콩잎에 에틸렌을 처리함으로써 이소플라본 함량을 높일 수 있다고 하였다.
본 연구에서는 콩의 발아기간 동안 몇 가지 물리적 처리를 통하여 이소플라본 함량의 증가 여부를 확인하였기에 이를 보고하고자 한다.
재료 및 방법
공시재료 및 콩 발아
국립식량과학원에서 2017년에 재배된 대풍2호, 우람 및 풍산나물콩 등 콩 3 품종을 공시재료로 사용하였다. 공시재료의 발아실험은 28°C 생육상에서 수행하였는데 페이퍼 타올에 100립씩 콩을 치상한 후 완전임의로 3반복으로 배치하고 하루에 4번 충분하게 수분을 공급하였다. 치상된 콩 종자는 발아 후 1일, 3일, 5일째에 각각 회수하여 이소플라본 분석에 사용하였다. 한편 5일 차 시료는 종자가 어느 정도 생육이 진전되었으므로 자엽과 배축을 따로 분리하여 각각의 이소플라본 함량을 분석하였으며, 1일 차와 3일 차 시료는 자엽부위만 분석에 사용되었다.
발아 중 물리적 요인 처리
열처리 및 초음파 처리가 이소플라본 함량에 미치는 영향을 알아보았다. 40°C에서 1시간 열처리 한 콩과 30분간 초음파(50 Hz)를 처리한 콩 각각을 대상으로 발아실험을 수행하고 일자별로 시료를 채취하여 동결건조한 후 이소플라본 함량을 분석하였다. 또한, 수분공급을 제한한 조건(대조군 대비-수분 공급량 1/2 처리)과 암 발아 대신 24시간 조명을 비추는 조건 각각에서 발아실험을 수행하였으며, 역시 일자별로 시료를 채취한 후 동결건조하고 이소플라본 함량을 분석하였다.
이소플라본 함량 분석
동결건조된 시료는 마쇄기로 100 mesh로 분쇄한 후 HPLC (Ultimate3000 HPLC, Thermo Dionex, USA)를 사용하여 이소플라본 함량을 분석하였다. HPLC 분석을 위한 전처리는 먼저 마쇄한 시료 0.1 g을 80% MeOH 30 ml로 vortexing한 후 30분간 sonication 하고 다시 180 rpm에서 1시간 shaking 하였으며, 이후 0.2 um PVDF로 filtering 하고 이소플라본 함량을 분석하였다. HPLC 분석에는 Inno C-18 column (4.6 x 250, 5 um, Youngjinbiochrom, Koera)과 UV-detector의 260 nm 파장을 사용하였다. 용매는 0.1% formic acid와 acetonitrile을 혼합하여 조합별로 gradient에 따라 1.0mL/min 속도로 흘려주었다. 이소플라본 표준물질은 12종은 모두 시중에서 구입하여 사용하였다. Aglycone 3종(glycitein, genistein, daidzein)과 glucoside 3종(glycitin, genistin, daidzin)은 Extrasynthese사(HPLC≥95%, France)의 제품을 사용하였고 malonyl-glucoside 3종(6’-O-malonyldaidzin, 6’-O-malonylgenistin, 6’-O-malonylglycitin)과 acetyl-glucoside 3종(6’-O-acetyldaidzin, 6’-O-acetylgenistin, 6’-O-acetylglycitin)은 Wako사(90%, Japan)의 제품을 사용하였다.
통계분석
모든 실험은 3반복으로 수행되었으며, 실험결과는 SPSS (Version 17.0, SPSS Institute Inc., Chicago, IL, USA)를 사용하여 통계처리 하였다.
결과 및 고찰
물리적 요인이 발아 중 이소플라본 함량에 미치는 영향
이소플라본 함량은 콩의 재배시기 동안 환경적 요인에 영향을 크게 받는데(Dhauhadel et al., 2003; Gutierrez-Gonzalez et al., 2010; Ahmad et al., 2017), 콩 종자에서도 이소플라본 축적이 환경의 영향을 받으며 자엽이 배축보다 환경에 더 민감하다는 연구결과가 있다(Berger et al., 2008). Kim et al. (1999)은 콩의 배아에 이소플라본이 많이 집적되어 있다고 하였다.
Table 1은 5일간의 콩 발아 기간 동안 몇 가지 물리적 요인에 의한 총 이소플라본 함량의 변화를 나타낸 것으로 분석 data는 공시된 3품종의 평균값이다. 총 이소플라본 함량은 물리적 요인에 상관없이 발아시간이 경과함에 따라 증가하는 경향이었다. 그런데 5일 차 자엽에서의 분석결과 일부가 발아 3일 차 보다 낮은 것은 5일 차 시료가 자엽과 배축으로 분리하여 이소플라본 함량이 분석되었기 때문이다. Chung (1998)과 Kim et al. (1999)은 콩에서 종자 발아 시 시간경과에 따라 이소플라본 함량이 증가된다고 하여 본 연구결과와 일치하였다. 그러나 콩의 이소플라본 함량은 발아 중반까지는 높아졌다가 다시 감소한다는 연구결과도 있다(Kim et al., 2004; Jeon et al., 2005). 한편, 배축의 이소플라본 함량이 다른 발아 일수에 비해 2배 이상 높았는데 이는 모든 결과 값이 시료의 건물중으로 분석되었기 때문이며 추후 이 결과 값을 활용하기 위해서는 생체중 혹은 무게 비율로 재조정하여 비교분석 되어져야 할 것이다.
Table 1. Total isoflavone content of soybean seeds subjected to abiotic treatment during the 5 germination periods. (ug/g, DW)
발아기간 중 처리된 물리적 요인 중 초음파 처리가 발아일수에 상관없이 이소플라본 함량이 가장 높았다. 특히 발아 3일 차 초음파 처리에서는 공시된 3개 품종의 평균 이소플라본 함량이 6,042.1 ug/g을 나타내어 원료콩의 이소플라본 함량인 4,609.5 ug/g(분석자료 미제시)보다 131% 증대됨을 알 수 있었다. 암 조건이 아닌 24시간 조명처리 조건에서의 이소플라본 함량은 평균값에서 초음파 처리와 유의성이 없었지만 품종 별 함량은 3 품종 모두 초음파 처리가 다소 높은 것으로 나타났다(Table 2). 발아 1일 및 3일 차 열처리에서도 이소플라본 함량이 초음파 처리와 유의성이 없었으나 품종 간에서는 차이가 나타났다. 그런데 열처리 된 시료는 공시 품종에 상관없이 발아기간 동안 부패립이 상당수 발견되었는데 이는 열처리가 콩의 발아에 좋지 않은 영향을 끼쳤기 때문으로 사료된다. 따라서 열처리가 이소플라본 함량을 높이는 기작에 영향을 미칠 수 있겠으나 전체적인 발아 상태로 판단할 때 열처리는 이소플라본 함량 증진 방법으로는 부적당하리라 판단된다. 한편, 발아 중 수분제한 조건은 대조군과 비교하더라도 이소플라본 함량이 낮았다.
Table 2. Total isoflavone content of soybean seeds subjected to abiotic treatments, arranged by cultivars and germination periods. (ug/g, DW)
Yang et al. (2015)은 초음파가 콩 발아 동안 glucoside (daidzin과 genistin)함량을 낮추는 대신 aglycone (daidzein과 genistein)함량은 높인다고 한 바 있어 일부는 본 연구 결과와 일치하기도 한다(Table 3 and Table 4). 또한, 초음파가 콩의 발아율을 증대시킬 뿐만 아니라 GABA 함량 증대, lipoxygenase activity 및 tripsin inhibitor 함량을 저하시켜 콩나물의 품질을 향상시키는 효과가 있다고 하였다. 그러나 그 기작에 대한 연구 결과는 아직까지 많이 부족하다. 이밖에도 콩의 발아 기간 동안 외부적 요인 처리가 콩 종자에 미치는 영향에 관한 연구들이 발표된 바 있다(Shohag et al., 2012; Yun et al., 2013).
Table 2는 공시품종별로 콩 발아기간 동안의 이소플라본 함량 변화를 나타낸 결과이다. 세 품종 모두 이소플라본 함량은 발아시간이 경과함에 따라 증가하는 경향이었다. 대풍2호의 경우 1일 차, 3일 차 및 자엽에서는 초음파와 조명 처리에서 이소플라본 함량이 가장 높았으며 다음으로 열처리 > 대조군 > 수분제한 순서로 함량에 차이가 있었다. 다만 초음파와 열처리는 자엽에서의 함량이 발아 3일 차 보다 낮아졌다. 우람에서는 발아일수에 상관없이 초음파 처리에서 가장 높았다. 다음 순서로는 함량간 유의성은 없었지만 열처리가 조명처리보다 높은 경향이었으며, 대조군 > 수분제한 순서로 이소플라본 함량이 높았다. 그러나 열처리는 앞에서 언급한 바와 같이 발아기간이 경과함에 따라 품종에 상관없이 부패립 발생이 문제점으로 나타났다. 풍산나물콩에서의 이소플라본 함량은 발아 1일, 3일 및 자엽에서 초음파와 조명처리가 이소플라본 함량을 높였으며, 기타 다른 처리들은 별 차이가 없었다. 그리고 모든 처리에서 자엽에서의 이소플라본 함량이 발아 3일 차 보다 떨어지는 것으로 나타났다.
발아 3일 차의 이소플라본 함량을 품종 별로 분석해 보면, 대풍2호가 모든 처리에서 다른 품종보다 이소플라본 함량이 높았으며 다음으로는 우람 > 풍산나물콩의 순서이었다. 그런데 대풍2호와 우람은 초음파 처리에서 가장 높고 수분제한 조건이 가장 떨어지는 등 처리별로 이소플라본 함량의 변화가 비슷한 경향을 보였지만 풍산나물콩은 처리간 차이가 별로 나타나지 않았다. 따라서 이소플라본 함량은 환경 및 유전적 요인 뿐만 아니라 환경과 유전적인 요인의 상호작용도 영향을 끼치리라 판단된다. Ahmad et al. (2017)은 여러 가지 스트레스 조건에서 이소플라본 함량을 분석한 결과 스트레스 종류에 따라 조직 및 기관에서 나타나는 이소플라본 함량에 차이가 있고 유전자의 발현도 다르다고 하였으며, Gutierrez-Gonzalez et al. (2009)은 주어진 환경에서 이소플라본 함량은 epistasis 등 유전자간 상호작용과 함께 유전적인 요인이 50% 이상을 차지한다고 하였다.
Fig. 1은 발아 5일이 경과한 후 배축에서의 이소플라본 함량을 물리적 요인과 함께 품종별로 나타낸 것이다. 배축의 이소플라본 함량은 Table 2의 발아 3일 차 자엽의 이소플라본 함량과는 다르게 나타났다. 즉 종자가 발아하는 동안 자엽과 배축에서의 이소플라본 축적은 양상이 다르다는 것을 알 수 있었다. 특히 풍산나물콩에서는 조명 > 초음파 > 수분제한 > 대조군 > 열처리 순서로 이소플라본 함량이 높게 나타나 배축에서도 이소플라본 축적 양상이 다른 품종과는 차이를 보였다.
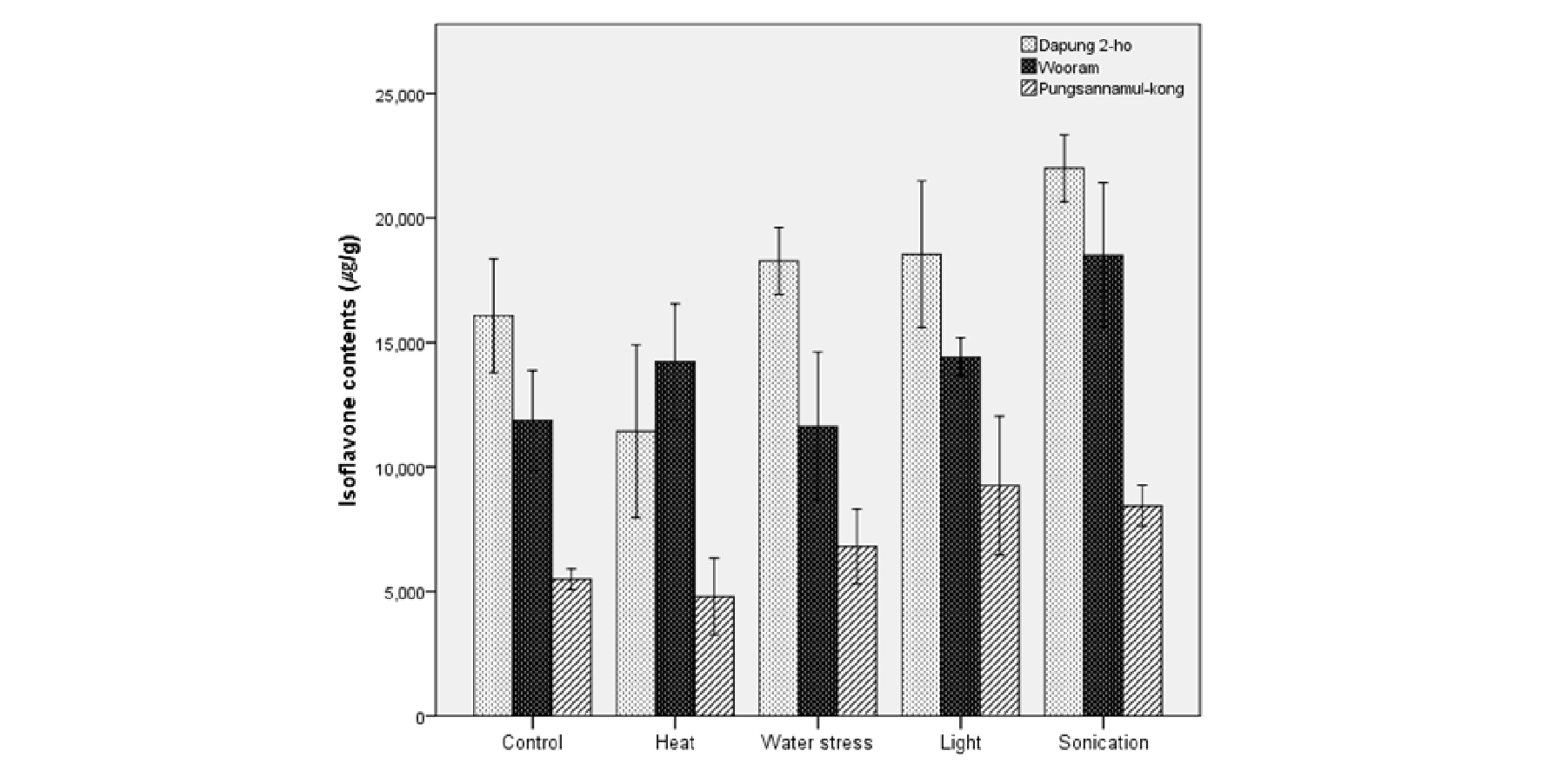
콩 발아와 이소플라본 종류별 함량 변화
콩의 이소플라본에는 aglycone 3종, glucoside 3종, malonyl-glucoside 3종 및 acetyl-glucoside 3종 등 12종이 있다. 그동안 이소플라본에 관한 연구는 주로 aglycone과 glucoside 중심으로 이루어져 왔는데 이는 malonyl-glucoside 및 acetyl-glucoside 함량 추적이 어려웠기 때문이다. 그러나 최근에는 전체 12종의 분석에 대한 연구결과가 있으며, 특히 콩 종자에 함유된 주된 이소플라본은 malonyl-glucoside이며 전체 이소플라본의 80% 이상을 차지한다고 한다(Ahmad et al., 2017; Lee et al., 2018).
Table 3은 콩 발아기간 동안 물리적 요인에 의한 이소플라본 group별 함량 변화를 나타낸 것이다. 이소플라본 종류별 함량을 분석한 결과 발아시간 경과에 상관없이 malonyl-glucoside > glucoside > aglycone > acetyl-glucoside의 순서로 함량이 높았다. 이중에서 malonyl-glucoside 함유량이 대체로 85% 이상으로 기존의 연구결과와 일치하였으며, aglycone과 acetyl-glucoside 함유량은 적었다.
Table 3. Content of each type of isoflavone group during germination of soybean seeds subjected to abiotic treatments. (ug/g, DW)
물리적 요인 처리에 따르면 초음파 처리와 열처리가 발아시간 경과에 상관없이 모든 이소플라본 종류에서 함량이 높았다. 한편, 이소플라본 함량은 이소플라본 종류별로 콩의 조직에 따라 축적 양상이 조금 다른 것으로 나타났다. 자엽의 경우, aglycone 함량은 초음파 처리 67.4 ug/g, 열처리 60.8 ug/g으로 대조군의 22.2 ug/g보다 3배 이상 높았으며, 이 함량은 발아 3일 차의 경우보다도 2배 가량 높았다. 또한, glucoside 함량도 초음파 처리 시 자엽에서 853.2 ug/g을 나타내어 대조군의 661.4 ug/g보다 높았다. 따라서 자엽에서의 이소플라본 축적 기작은 발아 1일 및 3일 차와는 다른 것으로 판단된다. 이것은 스트레스 종류에 따라 조직 및 기관에서 나타나는 이소플라본 함량에 차이가 있다는 보고(Ahmad et al., 2017)와 유사하였다. 이소플라본 축적에는 IFS 유전자의 발현이 중요한데(Yu et al., 2000), Gutierrez-Gonzalez et al. (2010)는 외부 스트레스에 상관없이 IFS1 유전자는 변함없이 발현되나 한발 조건에서는 IFS2 유전자의 발현에 차이가 나므로 스트레스 등의 환경조건에서는 IFS2 유전자의 발현 정도가 중요하다고 하였다.
한편, 인체에서는 이소플라본이 소화 후 혈장에서 약 24시간 존재하며, 평균적인 반감기는 6-8시간이라는 연구결과가 발표된 바 있으며(Manach et al., 2005), Yu et al. (2016)은 이소플라본 glucoside는 체내 소장에서 glucosidase에 의해 aglycone으로 가수분해되며, genistein이 daidzein보다 생리활성이 높다고 하였다. 최근에는 prenylated isoflavonoid의 생리활성 가능성이 제기되는 등(Yoneyama et al., 2016) 기존의 aglycone에 집중되었던 이소플라본 연구가 다양한 이소플라본 종류에 대한 연구로 확대되고 있다.
Table 4는 콩 발아기간 동안 물리적 요인에 의한 이소플라본 12종의 함량 변화에 대한 결과이다. Aglycone의 경우 발아 1, 3일 차 및 자엽에서 발아시간 경과에 따라 genistein은 계속 증가하는 반면 daizein과 glycitein 함량은 자엽에서 정체 혹은 감소하였다. 일반적으로 glycitein은 genistein과 daidzein에 비해 함량이 적었는데, 이는 aglycone 뿐만 아니라 glucoside 및 기타 glucoside에서도 같은 경향을 보여 콩에는 glycitein류의 함량이 적다고 판단할 수 있었다. 발아기간 및 품종에 따른 변화를 보면 daidzein은 우람 자엽에서 함량이 높았으며 glycitein은 대풍2호 3일차 및 우람 3일 차에서 높았다. Genistein은 우람 자엽에서 발아 1일 차에 비해 10배 가까이 높아졌으며, 대풍2호와 풍산나물콩의 자엽에서도 발아 1일 및 3일에 비해 상당히 높았다. Glucoside에서 daidzin의 경우 발아 3일차 및 자엽에서는 함량이 대동소이 하였으나 glycitin은 시간 경과에 따라 함량이 계속 감소하는 반면 genistin 함량은 계속 증가하는 경향이었다. Glucoside에서 이소플라본 종류별 축적 양상은 malonyl-glucoside에서도 같은 경향을 보였다. 품종간 비교를 하면 M-daidzin은 우람, M-glycitin과 M-genistin은 대풍2호에서 높았다. Acetyl-glucoside의 경우 A-glycitin은 분석되지 않았으며, A-daidzin과 A-genistin은 공시된 3 품종 공히 발아 3일 차 보다 자엽에서의 함량이 낮았다. 이는 발아 5일 차에 배축과 자엽으로 분리되어 함량이 분석된 영향이 있겠으나 acetyl-glucoside의 특성일 수도 있으리라 사료된다. 품종 및 발아기간별로 함량 차이를 보면 A-daidzin의 경우 대풍2호의 1일, 3일 및 자엽에서 높았으며, A-genistin은 대풍2호 3일 차에서 함량이 높았다. 한편, aglycone과 acetyl-glucoside에서는 daidzein류가 genistein류 보다 함량이 높았으나 glucoside와 malonyl-glucoside에서는 genistein류 > daidzein류 > glycitein류의 순서로 함량간 차이가 있었다.
Table 4. Content of each isoflavone type according to soybean cultivar and germination period. (ug/g, DW)
배축에서 이소플라본 종류별 함량은 Fig. 1에서 언급된 바와 같이 발아 1, 3일 차 및 자엽에서의 양상과는 달랐다. 특히 배축에서는 자엽에서 함량이 미미했던 glycitein류의 축적이 많이 이루어진 것을 볼 수 있었으며, glucoside와 malonyl-glucoside에서는 glycitin과 M-glycitin의 함량이 genistin과 M-genistin보다 높았다.
이상의 연구결과로 볼 때 콩 종자의 이소플라본은 발아를 통하여 그 함량을 높일 수 있으며, 나아가 발아 시 물리적 요인을 처리함으로써 특히 초음파 처리 등으로 이소플라본 함량을 한층 높일 수 있다고 판단된다.
적 요
콩 발아기간 중 몇 가지 물리적 요인(열 처리, 수분공급 제한, 조명 처리, 초음파 처리)이 이소플라본 함량 증대에 미치는 영향에 대하여 알아보았다.
1. 대조군과 함께 4가지 처리는 모두 발아시간이 경과함에 따라 이소플라본 함량이 높아졌다.
2. 초음파 처리가 이소플라본 함량 증대에 가장 큰 효과가 있었으며, 발아기간 중 조명 처리도 유의성이 인정되었다.
3. 열처리도 이소플라본 함량 증대에 유의적인 효과가 있었으나 발아기간 중 부패립이 많이 발생하는 문제가 있었으며, 수분제한은 대조군에 비해 이소플라본 함량이 떨어지는 경향이었다.
4. 이소플라본 종류별 함량을 분석한 결과 malonyl-glucoside가 대체로 총 이소플라본 함량의 85% 이상을 차지하였고 다음으로는 glucoside > aglycone > acetyl-glucoside 순서로 함량이 높았으며, 발아가 진행되면서 aglycone 함량이 증가하는 경향이었다.
이와 같은 결과를 볼 때 발아기간 중 외부의 물리적 요인 처리가 콩 종자의 이소플라본 함량을 증대시킴을 확인할 수 있었다.
