

서 론
벼는 단일성 작물로서 짧아지는 일장에 반응하여 출수를 한다(Bernier & Perilleux, 2005; Corbesier et al., 2007; Tamaki et al., 2007). 일장반응을 제외하면 온도에 따라 반응하며 평균기온 약 29~30°C까지는 온도가 높아질수록 출수가 촉진된다(Yoshida, 1981; Haniu et al., 1983; Loomis & Connor, 1992). 온도에 따라 출수가 촉진되는 정도는 품종에 따라 약간의 차이가 있지만 크게 다르지는 않다(Choi et al., 2002). 저온저항성 또는 고온저항성 등의 품종을 구분 짓는 특성은 생육적온 범위 밖에서 스트레스 반응을 유발하는 온도에서의 반응이며 벼의 생육과는 다른 개념이다(Satake & Yoshida, 1978). 같은 날 여러 품종을 이앙하였을 경우 품종마다 출수하는 시기가 같지 않은 이유는 품종들이 요구하는 한계일장이 달라 이에 반응하는 정도가 달라지기 때문이다(Collinson et al., 1992; Mimoto et al., 1989). 최적일장(Optimum photoperiod)이란 출수까지 최단 소요일수를 나타내는 일장을 말하며 한계일장(Critical photoperiod)이란 최장의 출수일수를 나타내거나 그 이상에서는 출수를 하지 않는 일장을 말한다.
일반적으로 벼의 출수 반응정도는 하루 중의 유효온도와 각 품종의 한계일장 보다 짧아진 일장조건에서의 감광반응과의 곱으로 표현하며, 어느 시기동안 이 값들이 누적되면 화아분화를 시작한다고 한다(Ahn, 1968; Collinson et al., 1992; Dua et al., 1990). 온도 측면에서 보면 벼가 생존할 수 있는 최저온도는 평균기온으로 약 10°C, 생육과 출수가 촉진되는 한계온도는 평균기온으로 약 30°C 내외이며(Vong & Murata, 1977; Best, 1959), 일장 측면에서는 10~12시간까지 일장이 짧아질수록 벼의 출수는 촉진된다(Choi et al., 2002). 온도와 일장 모두 벼의 출수에 크게 영향을 미치지만 어느 것이 더 영향력이 있는지에 대해서는 분명하게 밝힌 결과는 아직까지 보고되지 못하고 있다(Kovi et al., 2013; Bouman et al., 2001). 그러나 그 둘을 곱으로 표현하면 영향력의 차이는 소거된다. 하루의 평균기온이 25°C이며 일장이 14시간일 경우 그 날의 출수반응 정도는 (X(품종의 한계일장)−14(현재일장))/(X(품종의 한계일장)−12(최적일장))×(25(현재온도)−10(최저온도))/(30(최고온도)−10(최저온도))로 표현된다.
매년 같은 날의 일장의 변화는 차이가 없기 때문에 최근 농가에서 많이 재배되고 있는 품종들의 유효한계일장을 알게 되면 연차간 출수에 영향을 미치는 요인은 온도만 남게 되므로 온도변이에 따른 출수기 예측을 보다 쉽게 할 수 있다.
따라서 본 시험에서는 온도와 일장을 조절할 수 있는 인공기상실을 이용하여 벼의 출수반응을 연구하고 그 결과 따라 농가 재배 주요품종들의 한계일장을 추정한 결과를 제시하여 앞으로 농업현장에서 보다 정확한 출수기를 예측할 수 있도록 하고자 한다.
재료 및 방법
2016년도에 전국 벼 재배면적의 약 90%를 차지하였으며 2018년에도 재배면적의 약 85%를 차지하였던 농가재배 주요 벼 품종들에 대한 온도와 일장반응을 알아보고자 2017~ 2019년까지 3년간 국립식량과학원 인공기상실에서 시험을 실시하였다. 시험품종은 전체 18품종으로 조생종인 운광, 오대, 고시히카리, 조평 등 4품종, 중생종은 대보, 하이아미, 삼덕 등 3품종이며, 중만생종은 새누리, 신동진, 추청, 삼광, 일품, 새일미, 황금누리, 동진찰, 일미, 호품, 영호진미 등 11품종이었다. 온도 반응을 보기 위해서는 일장조건은 12:00으로 고정하였고 온도는 평균기온 22.0 (±5°C), 23.5 (±5°C), 25.0 (±5°C), 26.5 (±5°C), 28.0°C (±5°C) 5개의 처리로 벼를 재배하였다. 하루 중의 온도변화는 매시간 온도를 변화시켜 낮 최고기온은 평균기온보다 5°C 높고 아침 최저기온은 평균기온보다 5°C 낮게 설정하였다. 또한 일장반응 만을 보기 위해 처리온도는 평균기온 28°C (±5°C)로 동일하게 하였으며 최저 23°C, 최고 33°C로 설정하고 일평균기온 28°C인 날의 온도변화와 유사하게 매 시간대별로 온도를 변화시켰다. 일장처리는 12:00, 13:00, 13:10, 13:20, 13:30, 13:40, 13:50, 14:00, 14:10, 14:20, 14:30 등 11개 일장을 처리하였다.
재배방법은 1/5,000 와그너폿트에 발아된 종자 6개씩을 약 1 cm 깊이로 파종하는 직파재배법을 사용하였으며, 벼의 출아가 완료된 후 폿트당 3개체만 남기고 나머지는 제거하였다. 시설의 면적이 충분하지 않아 각 품종별 폿트 2반복(6개 식물체)으로 처리하였다. 기타 재배법은 농촌진흥청의 벼 표준재배방법에 준하였다.
출수기는 품종별 처리된 6개 식물체 중에서 가장 이삭이 먼저 나온 줄기를 기준으로 측정하였고, 이삭 추출 정도는 지엽에서 약 2/3정도 이삭이 추출된 시기를 출수의 기준으로 하였다. Table 2의 측정 데이터를 이용한 Fig. 2의 추정식은 일장이 길어지면 출수기도 지연된다는 것은 많은 연구를 통해 이미 충분히 알려진 사실이기 때문에(Katayama, 1980), 동일한 품종 내에서 보다 짧은 일장에서의 출수 소요일수가 보다 긴 일장에서의 출수 소요일수보다 긴 것으로 나타난 성적은 환경적 스트레스에 의한 출수 지연이라는 오류가 명확하여 추정식 생성을 위한 성적에서 제외하였다.
Table 3의 매일 발생될 처리 일장에 따른 감광반응 값은 처리별 일장요인(가장 좌측의 한계일장−처리일장)/(가장 좌측의 한계일장−최적 반응일장(12:00))과 온도요인인 (28−10)/ (29.2−10)의 곱을 표현한 값이다. 여기서 감광반응 값은 한계일장으로부터 최적일장으로 단일조건일수록 점점 더 커지는 값을 보인다. 온도요인의 분자인 28°C는 이 시험의 처리온도이고 분모인 29.2°C는 시험에 사용된 18개 품종의 한계온도이며 10°C는 벼의 생육 최저온도를 나타낸다. 온도요인은 동일하게 적용되었으므로 적용 온도가 달라질 경우에는 수치만 달라질 뿐 일장별 효과에는 영향을 미치지 못하기 때문에 품종별 동일한 일장반응 경향을 나타내기 위하여 18개 품종의 한계온도를 사용하였다. 표 우측의 화아분화 값은 처리일장에 따른 일별 감광 반응 값에 각 일장처리별 화아분화 시점까지의 소요기간(출수 소요일수−30)을 곱한 값으로 화아분화에 필요한 일별 감광반응 값의 누적을 나타낸다. 따라서 표 우측의 각 일장별 화아분화 값이 동일하게 되는 한계일장이 일품의 한계일장이 된다. 각 일장별 화아분화 값의 일치성 정도를 나타내기 위해 표의 가장 우측에 변동계수 CV (%) 값을 표시하였고 CV 값이 가장 낮은(2.07) 위치의 일장이 바로 화아분화 시점이 출수 전 30일로 가정된 일품의 한계일장이다. 한계일장의 CV (%)값과 CV (%)값이 가장 적은 지점의 값을 구할 때는 처리한 일장 이상 및 이하의 비교대상이 없어 오차의 우려가 매우 높은 처리일장 12:00과 14:30의 성적은 제외하고 나머지 성적들의 값을 이용하였다. 한계일장을 구하는 단계는 일품을 예시로 사용하였으며, 한계일장 및 화아분화 시점은 모든 품종의 값을 결과에 제시하였다.
결 과
온도에 따른 출수반응
온도에 따른 벼의 출수 반응의 일반적인 경향을 알아보기 위해 일장은 단일 조건인 12:00으로 고정하고 22°C에서 28°C까지 1.5°C 간격으로 벼를 재배한 결과는 Fig. 1에서 보는 바와 같으며, 시험에 사용된 18개 품종의 평균치로 보아 평균기온 1°C 상승에 따라 출수기간은 평균 약 3.5일 정도 단축되는 것으로 나타났으며, 수식을 분석한 결과 평균기온 29.2°C에서 출수일수의 최소값이 나오는 것으로 추정되었다. Table 1에는 각 품종별 온도에 따른 파종부터 출수까지의 일수를 나타내었다. 22°C 대비 28°C에서의 출수까지 생육일수의 차이는 조생종의 경우 19.5일 중생종은 22.7일, 중만생종은 20.9일의 차이를 보였으나, 생태형 간의 차이 대비 동일한 생태형 내 품종 간 편차가 더 커, 생태형별로 뚜렷한 온도반응의 차이를 보이지 못하였다. 품종별로는 22°C 대비 28°C에서의 출수까지 생육일수의 차이가 고시히카리에서는 18일(최저), 삼덕에서는 25일(최고)로 온도반응에 따른 생육 단축 정도가 최대 약 7일 정도 차이가 나는 것을 확인하였다. 한편 22°C 와 25°C 처리구에서 생육일수의 차이가 18개 품종 평균 14.9일 이었으며, 25°C 와 28°C 처리구에서는 6.0일로 동일한 3°C 온도 차이지만 온도가 상승할수록(고온 한계온도에 가까워질수록), 온도에 따른 반응 차이가 작아지는 것을 확인할 수 있었다.
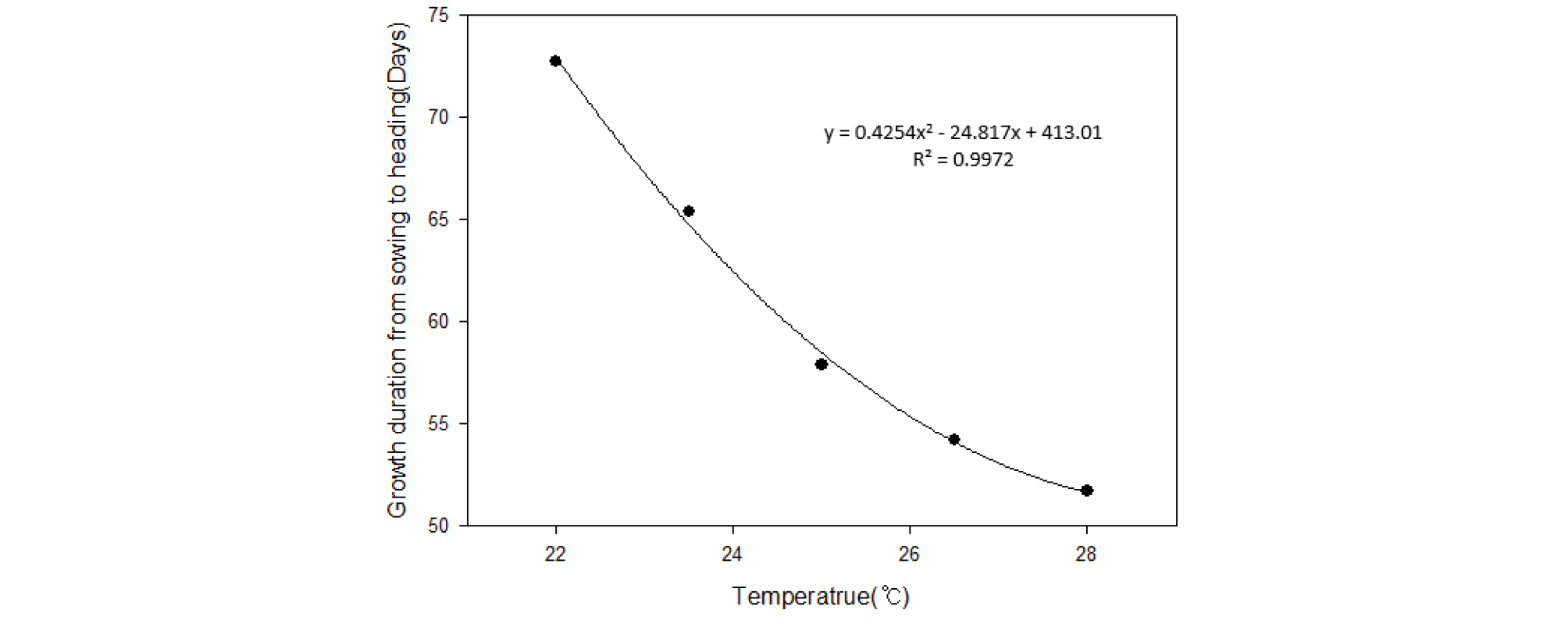
Table 1. Growth duration from sowing to heading by varieties in each different temperature conditions.
일장에 따른 출수반응
파종 후 처리일장에 따른 시험 품종들의 출수 소요일수를 조사한 결과는 Table 2에서 나타난 바와 같으며, 처리 일장 내 출수를 가장 촉진시키는 12:00 일장조건에서 파종~출수까지 소요일수는 품종에 따라 43~56일이었으며 14:30 일장에서는 영호진미를 제외한 모든 품종에서 61~75일 소요되었다. 영호진미는 14:20 일장에서 79일을 나타내었고 14:30 일장에서는 출수가 지나치게 지연되어 시험을 중단하였다. 품종 생태형으로 분류하여 본 12:00 일장과 14:30 일장과의 출수 소요일수 차이는 조생종 약 9.5일(8~13일), 중생종 약 18.3일(17~20일), 영호진미를 제외한 중만생종은 약 23.5일(16~29일) 소요되어 생태형별로 조생종과 중, 중만생종 간 뚜렷한 반응 차이를 보였다.
Table 2. Growth duration from sowing to heading under mean temperature 28°C (±5°C) in each different day-length.
Table 2에서 나타낸 일장별 출수 소요일수를 기준으로 각 품종별 일장에 따른 출수 소요일수에서 출수 지연에 따른 비정상적인 성적을 제거한 후 정상적으로 출수하였던 성적을 토대로 계산된 추정식을 나타낸 결과는 Fig. 2에서 보는 바와 같다. 각 그림에서 검은색 사각형으로 표시한 것은 출수 지연으로 제거된 오류를 나타낸 것이며 추정식 산정에 사용된 성적은 흰색 사각형으로 표시하였고 그림 하단에는 수식을 나타내었다. 일장 조건 별로 실험조건 내에서 비교적 장일 조건인 14:30에서와 13:30에서의 생육일수 차이는 생태형별로 조생종, 중생종, 중만생종 각각 6.5일, 11.4일, 13.6일 이였으며, 비교적 단일 조건인 13:00과 12:00에서의 차이는 생태형별로 조생종, 중생종, 중만생종 각각 0.7일, 4.4일, 4.7일로 같은 1시간 일장 차이에서도 일장조건이 짧아질수록 일장 반응에 따른 차이가 감소하는 것을 확인하였다.
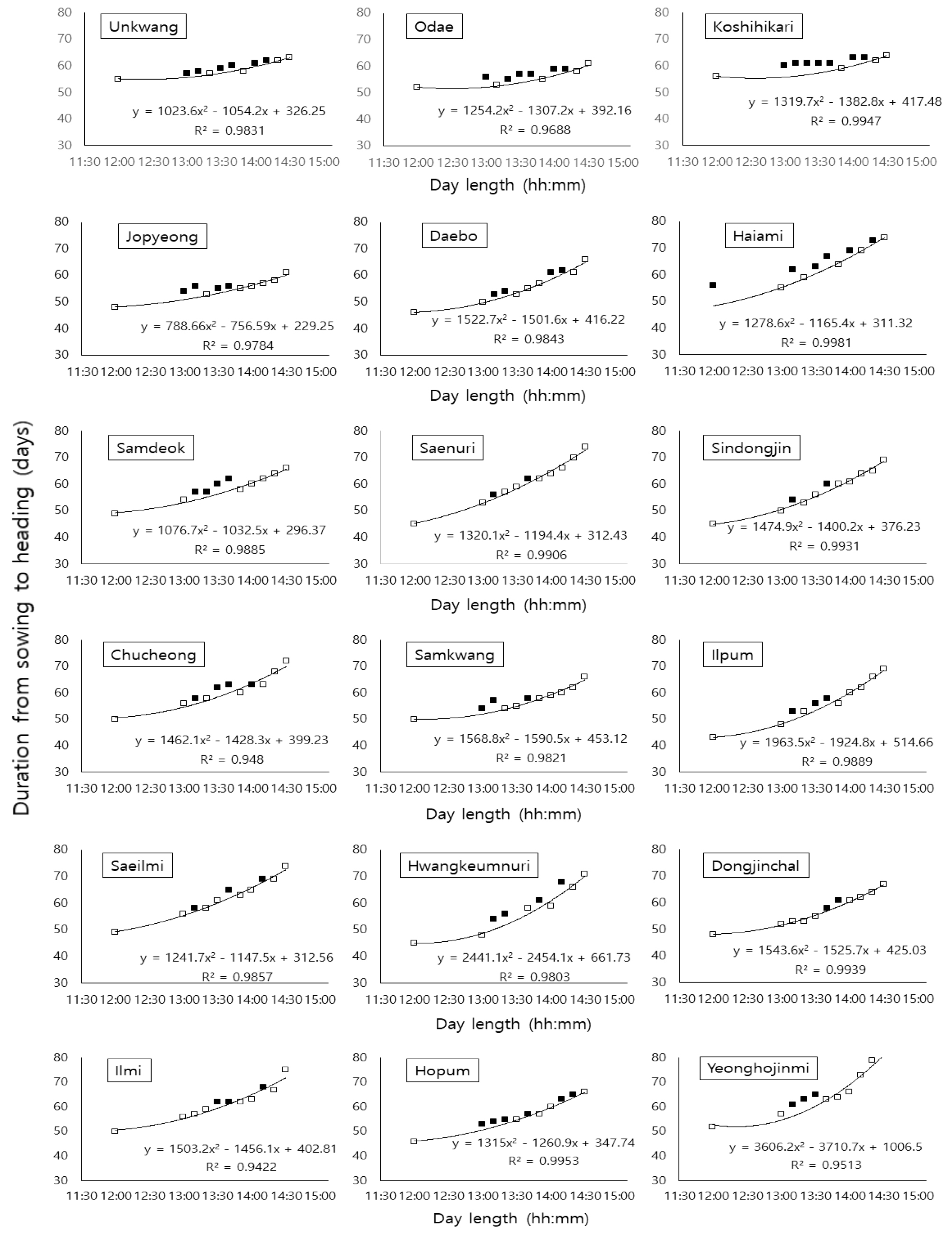
벼 품종별 한계일장
본 시험에서는 온도를 평균기온 28°C로 고정하여 각 품종들의 화아분화에 영향하는 일장들을 알아보았다. Fig. 2에서 계산된 일품의 각 일장별 출수 소요일수에서 화아분화 시점인 30 (출수 전 30일)을 제하면 파종부터 화아분화까지 필요한 소요기간은 일장 12:00에서 13.1일, 13:00에서 18.2일, 그리고 14:30에서는 38.5일 등으로 표시된다(Table 3). Table 3의 가장 좌측의 일장은 일품의 한계일장을 가정하여 나타낸 것으로 한계일장 23:50부터 15:39까지 나열하였고, 표 좌측에는 각 처리 일장별 감광반응 값, 표 우측에는 감광반응 값에 화아분화까지의 소요일수를 곱한 값인 화아분화 값을 나타내었다. 여기서 화아분화 값이 균일하게 나타나는 (CV 값이 가장 낮은) 한계일장을 분석하였다. 그 결과 일품의 한계일장은 15:44로 나타났고, 한계일장이 15:44으로 설정되었을 경우 화아분화 시점까지 최소한으로 필요한 화아분화 값은 12.5로 추정되었다.
Table 3. Estimating the critical day-length based on the flowering initiation is 30 days before heading in Ilpum variety.
*In the case of Ilpum variety, the minimum CV (%, except 12:00 and 14:50 day length) was appeared in 15:44 day-length
(B) : (23:40‑13:20)/(23:40‑12:00)×(28‑10/29.2‑10),
(C) : (B)×21.3(flower initiation data under 13:20 day-length),
(D) : minimum data among day-length(except 12:00 and 14:50) treatment
일품을 기준으로 화아분화 시점을 출수 전 30일로 가정하여 산출한 한계일장 15:44를 적용한 결과는 Table 4에서 보는 바와 같으며, 화아분화 시점까지 최소한으로 필요한 화아분화 값이 12.5 이상이 되는 최초의 값을 굵은 숫자로 표시하였다. 평균기온 28°C 조건에서 12:00 일장에서는 화아분화 시점이 파종 후 16일, 13:00 일장에서는 파종 후 21일, 14:00 일장에서는 파종 후 31일, 그리고 14:30 일장에서는 파종 후 43일로 나타나 출수까지의 소요일수는 각각 30일을 더한 값인 46, 51, 61, 그리고 73일로 추정되었다.
Table 4. Flower bud differentiation (30 days before heading) value (>12.5) in each day length treatment were displayed bold & underlined in the case of critical day length 15:44 in Ilpum variety.
Table 4에서 나타낸 방법과 같이 화아분화 시점까지 최소한으로 필요한 화아분화 값을 일품의 화아분화 시점이 달라질 경우를 가정하여, 각 일장별 출수 소요일수와 한계일장 및 화아분화 값은 Table 5에서 나타낸 바와 같다. 화아분화 시점을 출수 전 30~33일로 가정하였을 경우, 화아분화 시점이 출수 전 30일이었을 경우의 한계일장 15:44과 누적 값 12.5는 화아분화 시점이 출수 전 33일로 앞당겨지면 한계일장은 15:31, 누적 값은 10.2으로 변하여 화아분화가 빨라질수록 한계일장은 짧아지고 화아분화 값은 감소하였다.
Table 5. Changes of estimating growth duration from sowing to heading at different day-length treatment and critical day-length for heading based on the different flowering bud differentiation time in Ilpum variety.
*Estimating growth duration was calculated for each flower bud differentiation time (days after sowing) when the flower bud differentiation value reached the minimum value (12.5) as shown in Table 4
위와 같은 방법에 의해 화아분화 시점을 달리한 각 품종들의 한계일장과 누적 값을 표시한 결과는 Table 6에서 나타낸 바와 같다. 모든 품종의 한계일장과 누적 값은 일품과 동일한 경향을 나타내어 화아분화 시점이 앞당겨질수록 일정한 경향으로 한계일장은 길어지고 누적 값은 감소하였다.
Table 6. Critical day length and flower bud differentiation value for each varieties according to the time of flower bud differentiation time.
정확한 화아분화 시점을 찾아내기 위해 18품종의 평균치로 Table 4의 추정방법에 의한 각 품종의 일장별 파종부터 출수까지 소요일수에서 Fig. 2의 일장별 출수 소요일수를 뺀 결과는 Table 7에서 보는 바와 같으며, 각 일장별 차이를 평균하면 화아분화 시점이 출수 전 33일로 가정한 경우에는 평균오차가 1.0일로 나타나 시험성적 대비 1일 이른 것으로 나타났으며, 32일로 가정한 경우에는 0.0일로 시험성적과 유사하였고, 31일과 30일로 가정한 경우는 각각 -1.1과 -2.1일로 시험성적 대비 각각 1일과 2일 빠른 것으로 나타나 28°C 조건에서는 화아분화 시점을 출수 전 32일로 보는 것이 가장 타당하였다.
Table 7. Difference between recalculated growth duration and estimating growth duration according to flower bud differentiation time (mean of 18 varieties).
*Estimating growth duration data was calculated for each flower bud differentiation time (days after sowing) when the flower bud differentiation value reached the minimum value as shown in Table 4
따라서 농가재배 주요 품종들의 한계일장은 출수 전 32일을 화아분화 시점으로 계산된 결과를 적용하는 것이 바람직하며, 출수 전 32일을 화아분화 시점으로 계산하였을 경우 한계일장은 조생종인 운광에서 19:20, 오대 18:14, 고시히카리는 18:58, 조평은 17:30이었다. 중생종인 대보, 하이아미, 삼덕은 각각 16:08, 16:15 및 16:55이었고 중만생종인 새누리, 신동진, 추청, 삼광, 일품, 새일미, 황금누리, 동진찰, 일미, 호품, 영호진미는 각각 15:58, 15:56, 16:36, 16:44, 15:35, 16:26, 15:33, 16:20, 16:29, 16:13, 15:41이었다(Table 6).
일품을 기준으로 화아분화 시점을 출수 전 32일로 가정하여 산출한 한계일장 15:44를 적용한 결과는 Table 8에서 보는 바와 같으며, 화아분화 시점까지 최소한으로 필요한 화아분화 값이 10.9 이상이 되는 최초의 값을 굵은 숫자로 표시하였다. 최종적으로 화아분화 시점을 32일로 기준으로 Table 8과 같이 각 품종별 한계일장 및 화아분화 값을 이용하여, 일장조건별 출수소요일수를 예상한 결과는 Table 9에 나타낸 바와 같다.
Table 8. Flower bud differentiation (32 days before heading) value (>10.9) in each daylength treatment were displayed bold & underlined in the case of effective daylength 15:35 in Ilpum variety.
Table 9. Growth duration from sowing to heading under mean temperature 28°C (±5°C) in each different day-length.
고 찰
벼의 출수반응에 영향을 주는 큰 두 가지 환경요인은 일장과 온도이다(Ahn, 1968; Collinson et al., 1992; Dua et al., 1990). 출수까지의 벼의 생육기간은 Vergara et al. (1985)과 Roberts et al. (1987)은 발아부터 출수기까지를 일장에 감응하지 않는 기간과 일장에 감응하는 기간, 그리고 다시 일장에 감응하지 않는 기간으로 3 단계로 나누었다. 이는 Yin et al. (1997)의 실험에서 편의상 각각 Basic Vegetative Period (BVP), Photo Sensitive Period (PSP), Post Photo- sensitive Period (PPP)로 명명하였다. 온도와 일장에 대한 그간의 보고는 일장은 오직 PSP 기간에만 영향을 미친다거나, BVP기간을 제외한 PPP기간에도 영향을 미칠 수 있다는 상반된 의견이 존재하지만(Collinson et al., 1992; Mimoto et al., 1989), 온도는 전 생육기간에 영향을 미치며(Yoshida, 1981), 생육단계별로 출수에 미치는 영향력이 다르다고 하였다(Wang, 1960; Yin et al., 1997; Lee et al., 2019). 온도에 따른 반응은 평균기온 약 29~30°C까지는 온도가 높아질수록 출수가 촉진된다고 하였는데(Yoshida, 1981; Haniu et al., 1983; Loomis & Connor, 1992), 본 실험의 결과에서도 마찬가지로 평균기온 28°C까지 모든 품종에서 출수까지의 소요일수가 단축된 것을 확인하였으며, 생태형별로 온도반응에 따른 뚜렷한 차이는 보이지 않았다(Fig. 1, Table 1). 한편 온도에 따른 반응은 처리온도가 높아질수록 온도증가에 따른 출수까지의 소요일수가 단축되는 반응이 작아지며 한계온도 이상에서는 출수반응이 오히려 저해되었던 기존의 연구 결과들(Yin et al., 1996; Noguchi, 1959)과 마찬가지로, 본 실험의 국내 시험 품종에서도 29.2°C 까지 처리온도가 높아질수록 온도에 대한 출수까지의 소요일수 단축정도가 작아지는 것을 확인하였다(Table 1). 이러한 결과는 유수형성 반응에 주요한 조절 유전자인 Hd3a는 27°C 대비 23°C 처리조건에서 발현이 줄었지만, 상위유전자인 Hd1은 23°C, 27°C 조건에서 발현차이가 없었던 기존의 연구 결과를 통해(Luan et al., 2009), 유수형성 반응의 중 Hd3a 외 MADS14, MADS15의 발현을 조절하는 RFT1과 같은 다른 상위 유전자는 생육적온의 범위에서는 유수형성 반응에 온도에 따른 영향이 거의 없을 것으로 유추해 볼 수 있으며, 이에 대한 면밀한 검토가 필요할 것으로 보인다.
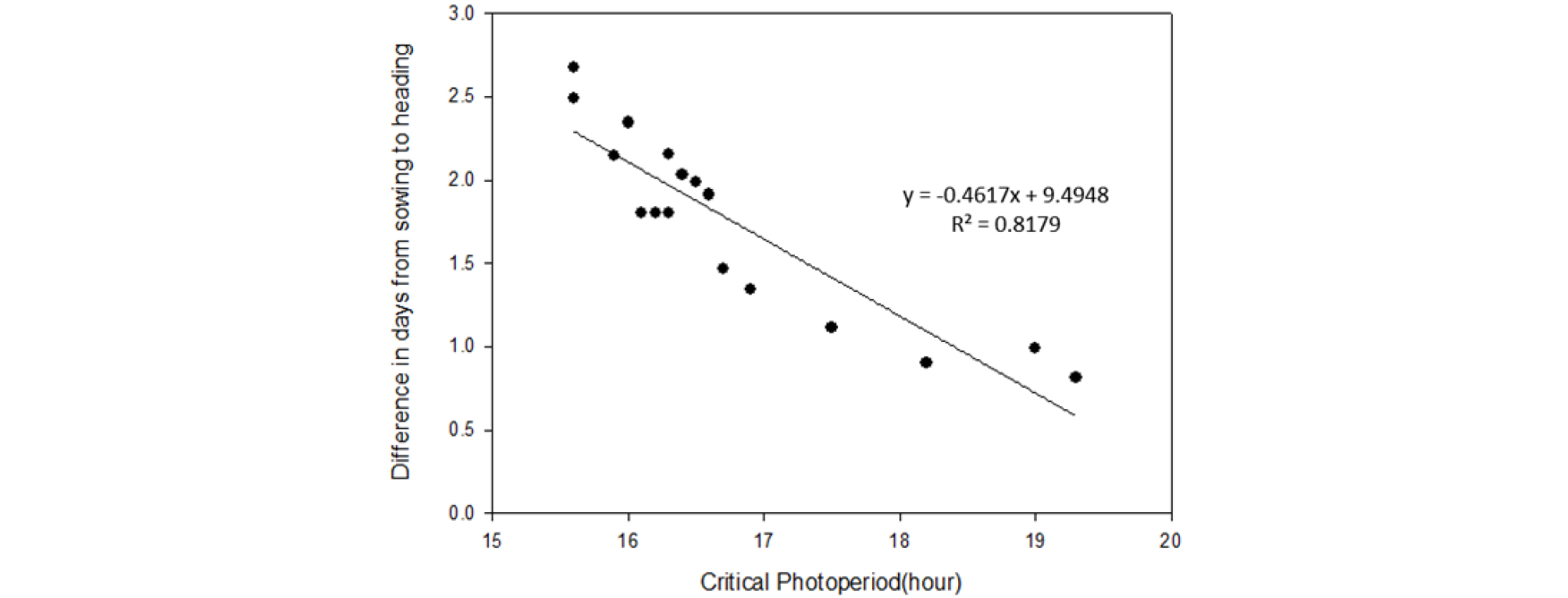
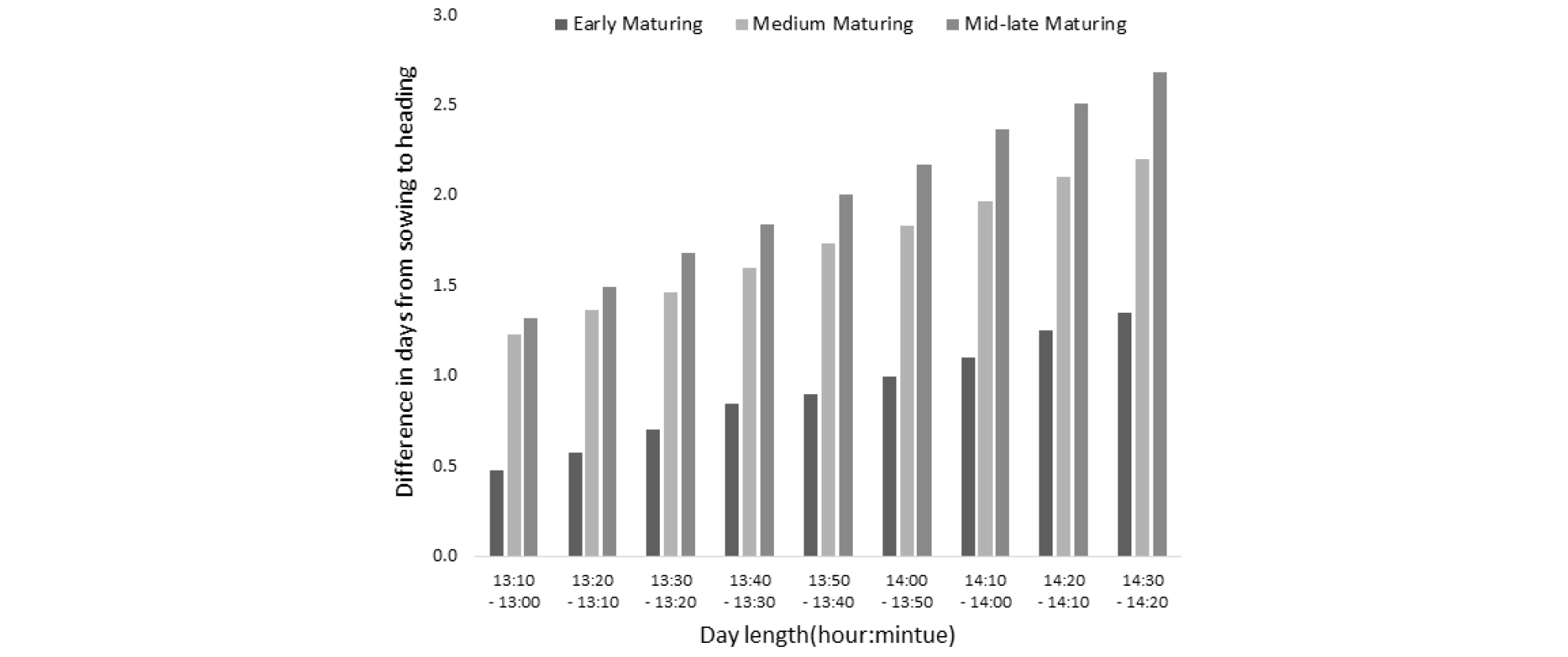
벼는 일장에 민감하여 장일 조건에서는 출수가 지연되며, 품종별로 일장의 민감성 정도에 대한 다양한 변이가 존재한다(Enomoto, 1935; Liang & Liu 1983; Morinaga & Kuriyama, 1954). 파종 후 처리일장에 따른 출수까지 소요일수는 온도 반응과는 달리 생태형별간 뚜렷한 반응 차이를 보였으며, 중만생종 품종에서 반응이 가장 큰 것으로 확인되었다(Table 2). 농가 재배 주요 품종의 한계일장 구명은 처리일장에 따른 일별 감광반응 값이 누적된(일별 감광반응 값에 각 처리 일장조건에서 파종부터 화아분화까지의 소요일수를 곱하여 계산) 개념인 화아분화 값을 설정하여 계산하였다(Table 3). 서로 다른 일장처리 조건에서 화아분화 값이 일치하는 일장이 바로 그 품종의 한계일장이 된다(Bouman et al., 2001). 화아분화 시점을 32일로 기준할 경우 한계일장은 조생종은 17:30에서 19:20으로 다소 길었고, 중생종·중만생종은 15:41에서 16:55로 조생종 대비 짧았는데, 이러한 생태형별 한계일장은 Fig. 3에서 보는바와 같이 각 품종의 14:30에서 13:00까지 10분 간격의 일장차이에 따른 평균 출수까지의 소요일수 차이, 즉 감광반응 정도와 음의 상관관계를 보이는 것을 알 수 있다. 결국 벼가 생육되는 일장조건에서는 한계일장이 짧은 품종일수록 각 품종의 한계일장 보다 짧은 일장 조건에서는 더 민감하게 반응 한다는 것을 알 수 있다. 또한 생태형별 감광반응 차이의 발생이 조생종은 감광을 할 수 있는 기간자체가 중만생종에 대비 짧기 때문으로 생각 할 수 있는데, 이는 한계일장보다 최적일장에 가까워질수록 감광반응을 할 수 있는 상대적으로 능력이 포화가 되어 일장차이에 따른 소요일수 차이가 작아지는데(Fig. 4), 국내에서 벼가 재배되는 시기의 일장은 조생종의 경우 한계일장보다 최적일장에 더 가깝지만, 중만생종의 경우 한계일장에 더 가깝기 때문에 반응 할 수 있는 기간 자체가 길어지는 것으로 생각된다. 하여 한계일장이 상대적으로 짧은 일품(15:35), 황금누리(15:33) 및 영호진미(15:41) 등의 품종은 이른 시기에 모내기를 하더라도 하지를 중심으로 한 긴 일장에서는 감응도가 매우 낮을 것으로 예측되어 이른모내기를 할 경우 출수일수를 앞당기기는 어렵고, 이앙이 매우 늦어질 경우 일장이 짧아진 시기에 반응성이 커서 늦어진 이앙시기 대비 출수기는 크게 앞당겨질 것으로 생각된다. 일장의 변화는 매년 차이가 없기 때문에 농가에서 많이 재배되고 있는 품종들의 유효한계일장을 알게 되면 연차간 출수에 영향을 미치는 요인은 온도만 남게 되므로 온도변이에 따른 출수기 예측을 보다 쉽게 할 수 있을 것으로 생각된다.
이러한 한계일장의 구분은 Itoh & Izawa (2013)는 Florigen gene인 Hd3a, RFT1 및 Repressor인 Ghd7의 발현으로 구분하였는데, 한계일장 이상의 장일조건에서는 Ghd7이 Hd3a의 발현을 억제하여, RFT1이 감광반응을 조절하여 반응 정도가 아주 낮지만, 한계일장 이하의 단일조건에서는 Hd3a가 RFT1과 함께 감광반응을 조절하여 반응 정도가 커진다고 하였다. 추후의 시험에서는 본 시험에서 구분된 각 품종의 한계일장에 따라서 실제로 Hd3a의 발현 특성 변화와 일장이 짧아져 최적일장에 가까워질수록 Hd3a와 RFT1의 발현 특성이 어떻게 변이되는지에 대한 분자생물학 수준에서의 검토가 필요할 것으로 생각된다.
적 요
벼 주요재배 품종의 온도 및 일장에 따른 출수반응을 구명한 내용은 다음과 같다.
1. 온도에 따른 출수반응은 일장 12:00, 평균기온 22°C에서 28°C까지의 범위에서 1°C 상승에 따라 평균 약 3.5일 정도 단축 되는 것을 확인하였으며, 온도가 상승 할수록 그 차이는 감소하는 경향을 보였다. 또한 온도에 따른 출수까지의 소요일수는 생태형별로 뚜렷한 차이를 보이지 못하였다.
2. 14:30과 12:00의 일장조건에 따른 출수까지의 소요일수 차이는 조생종 9.5일, 중생종 18.3일, 중만생종 23.5일로 온도반응과 달리 생태형간 뚜렷한 반응 차이를 보였으며, 일장조건이 짧아질수록 모든 생태형에서 일장반응에 따른 차이가 감소하는 것을 확인하였다.
3. 벼 주요 재배 품종의 화아분화 시점은 32일이었으며, 각 품종별 한계일장은 조생종인 운광, 오대, 고시히카리, 조평은 각각 19:20, 18:14, 18:58, 17:30이었다. 중생종인 대보, 하이아미, 삼덕은 각각 16:08, 16:15 및 16:55이었고, 중만생종인 새누리, 신동진, 추청, 삼광, 일품, 새일미, 황금누리, 동진찰, 일미, 호품, 영호진미는 각각 15:58, 15:56, 16:36, 16:44, 15:35, 16:26, 15:33, 16:20, 16:29, 16:13, 15:41이었다.
