

서 론
재료 및 방법
집단 육성
내탈립성 검정
생육특성 조사
내탈립성 유망계통 선발
통계 분석
결과 및 고찰
건조 시간에 따른 내탈립성 변이
연차별 내탈립성 변이
처리 시간에 따른 효과와 계통 간 내탈립성 변이
생육특성 조사 결과
내탈립성 유망계통 선발
적 요
서 론
콩의 성숙기 탈립은 식물의 자연스러운 종자번식의 한 형태로서 다양한 환경에 적응하기 위한 식물체의 기작으로 생각되며(Funatsuki et al., 2008), 특히 야생콩(Glycine soja Sieb. et Zucc.)의 경우 탈립에 매우 취약한 특성을 나타낸다(Suzuki et al., 2010; Funatsuki et al., 2014). 하지만 작물로서 콩이 재배되기 시작하면서 이러한 성숙기 탈립 문제는 종자 수확 시에 심각한 수량손실의 원인이며, 수확작업의 제한요인이 되어 입모관리와 함께 중요한 콩 재배기피 원인이 되고 있다. 최근에는 농촌 인구 고령화와 농업 인구수 감소로 노동력 절감을 위한 기계화 재배에 맞는 콩 품종이 요구되면서 콩의 성숙기 탈립 문제는 식물체의 쓰러짐과 함께 가장 해결되어야 할 불량 형질로 간주되고 있다.
식물체의 탈립 현상은 꼬투리가 달리는 작물인 두과작물이나 십자화과작물에서 발견된다. Arabidopsis thaliana에서는 탈립 저항성인 돌연변이체를 통하여 유전연구가 수행되었고, 이에 따라 성숙기에 식물체의 꼬투리 이층의 결합부를 강화하여 탈립에 저항성이 나타난다는 것이 알려졌다(Liljegren et al., 2000; Mitsuda et al., 2008; Ogawa et al., 2009). 콩에서는 야생콩으로부터 콩이 작물로서 재배되기 시작하면서 수확기에 식물체에서 탈립 되지 않은 개체가 선발되어 이듬해에 종자로서 활용 되는 식의 자연 선발과정이 진행 되었고, 이에 따라 재배종 콩은 야생콩에 비해 성숙기 탈립에 저항성인 방향으로 유전되어 왔을 것으로 추정하고 있다(Hancock, 2004; Fuller, 2007).
‘대원콩’(Kim et al., 1998a)은 1997년에 국립식량과학원에서 개발한 품종으로 수량이 낮고 쓰러짐이나 불마름병 등 병해에 취약한 특성을 가지고 있으면서도 성숙기 탈립에 매우 강한 특성을 나타내어 현재까지도 우리나라 가장 널리 보급되고 있는 우량품종이다. 따라서 향후 콩 육종 프로그램은 ‘대원콩’ 수준의 탈립성을 가지면서 수량성, 내병성, 품질 등의 형질이 보완된 품종을 개발하여 ‘대원콩’을 대체하는 것을 목표로 진행될 예정이다. 성숙기가 빠른 단기성콩, 재래종 검정콩 등은 특히 탈립에 매우 취약한 특성을 나타내는데, ‘대원콩’의 내탈립성에 대한 유전적 해석을 통하여 ‘대원콩’ 수준의 내탈립성을 갖는 유색콩, 단기성 콩 등의 중간모본 개발로 수량 손실을 최소화 할 수 있을 것으로 기대된다.
현재까지 국내 콩 품종개발에서 내탈립성은 그 중요성이 인정되면서도 초기세대에서 명확한 유전적 지식 없이 육종가의 판단으로 선발해왔다. 콩의 탈립성에 대한 QTL 분석 등 유전연구는 오래전부터 진행되어 왔고(Bailey et al., 1997; Funastsuki et al., 2006; Funatsuki et al., 2008; Kang et al., 2008) 이에 따라 소수의 유전자로부터 저항성이 유전된다는 것이 밝혀졌다. 2010년 ‘Williams82’ 품종에 대한 콩의 표준 유전체 해독이 완료된 이후로(Schmutz et al., 2010), SNP칩 개발, NGS분석 등 유전 분석 기술이 높은 수준으로 발달하게 되었고, 콩의 내탈립성에 대한 유전 연구가 급속히 진전되었다. 이 결과로 콩의 탈립성이 성숙기 건조 시에 꼬투리의 과도한 목질화에 의해 나타나며, 16번 염색체 상에 존재하는 SHAT1-5 유전자가 2세포벽 생합성을 활성화하여 꼬투리 봉합부 결합력을 향상시켜 내탈립성을 강화하고(Dong et al., 2014), pdh1 유전자가 건조 시 콩 꼬투리의 비틀림을 유도하여 탈립을 촉진시키는 것이 밝혀졌다(Funatsuki et al., 2014).
국내에서는 내탈립성 품종인 ‘신팔달콩’(Hong et al., 1992)을 부본으로 하는 집단을 대상으로 내탈립성 관련 유전분석이 수행되어 ‘신팔달콩’의 내탈립성이 pdh1 유전자에 의한 것으로 밝혀졌고, 해당 RIL 집단에 특이적인 SNP마커를 개발하기도 하였으나(Lee et al., 2017), ‘대원콩’ 유래의 내탈립성 연구는 아직까지 수행되지 않았다. 따라서, 본 연구의 목적은 ‘대원콩’을 모본으로 한 고세대 RIL 집단을 육성하여 집단 내에 내탈립성 변이를 조사하고, ‘대원콩’ 수준의 내탈립 특성을 가지는 유색콩, 단기성콩 유망계통을 선발하고자 하는 것이었다.
재료 및 방법
집단 육성
내탈립성 품종인 ‘대원콩’을 모본으로 하고 감수성인 검정콩 품종 ‘다원콩’(Kim et al., 1998b)과 단기성 콩인 ‘새올’(Baek et al., 1998)을 부본으로 인공교배를 실시하여 두 조합의 F1, F2 집단을 육성하였다. F3 이후에는 SSD로 전개하여 세대를 진전시켰으며, ‘16년부터’ 18년까지 연차별로 F6, F7, F8 세대의 재조합자식계통(recombinant inbred lines, RILs) 집단을 육성하였다. 육성된 두 조합의 RIL집단에서 3년간 연차별로 각 집단의 모·부본 및 ‘대원콩’ב다원콩’ 조합에서 154계통, ‘대원콩’ב새올’ 조합에서 153계통에 대하여 실내 내탈립성 검정을 수행하였다.
내탈립성 검정
2016년부터 2018년까지 3년간 두 집단의 모·부본 및 전체 307계통에 대한 실내 내탈립성 검정을 수행하였다. 시험구는 완전임의배치법 1반복으로 배치하였으며, 각 계통에서 성숙기(RDA, 2012)에 성숙된 협을 2반복으로 채취하여 상온에서 일주일간 치상 후 성숙기를 일치시켰다. 상온 치상 후 40°C 건조기에 건조하여 24시간, 48시간, 72시간 후 개열한 협의 비율(개열한 협의 수/채취한 협의 수×100%)을 구하였고(Kang et al., 2005), 1년차에는 성숙된 협 5개, 2, 3년차에는 성숙된 협 10개를 각각 2반복으로 채취하여 동일한 방법으로 실험을 수행하였다.
생육특성 조사
각 집단의 모·부본 및 ’16-’17년도 실내 내탈립성 검정에서 개열된 협의 비율이 0%로 조사되었던 내탈립성 71계통에 대하여 작물학적특성을 조사하였다. 생육조사는 국립식량과학원 남부작물부 내의 콩 시험 포장(경상남도 밀양시, 위도 35.5, 경도 128.7)에서 수행하였고, 시험구는 완전임의배치법 1반복으로 배치하였다. 계통 당 5개의 개체에서 생육조사를 수행하였으며, 생육특성은 농업과학기술 연구조사분석기준(RDA, 2012)에 따라 개화기, 경장, 착협고, 마디수, 분지수, 협수 등을 조사하였다. 재배방법은 재식거리 이랑 간 간격 70 cm,주 간 간격 15 cm, 1주 2본으로 재배하였고, 비료는 N:P:K=3:3:3.4에 준하였고, 퇴비는 10 a 당 1톤을 시용하였다.
내탈립성 유망계통 선발
3년간 수행한 실내 내탈립성 검정에서 건조기에서 72시간 건조 후 개열한 협의 비율이 0%인 계통을 선발한 후 생육특성 조사 결과와 종합하여 내탈립성 유망계통을 선발하였다. 각 집단에서 ‘대원콩’ 수준의 내탈립성을 가지면서 종피색이 유색인 계통과 개화기 및 성숙기가 빠른 단기성 계통을 선발하였다.
통계 분석
건조 시간대별 집단 간 탈립률 상호비교를 위해 각 집단 평균에 대한 t-검정을 수행하였고, 유전형 및 연차 반복이 탈립률 변이에 영향을 미친 유의성을 확인하기 위하여 연차별 전체 307계통의 분산분석(analysis of variance)과 건조 시간별, 연차별 탈립률을 통한 상관분석(correlation analysis)을 수행하였다. 분산분석 결과를 통하여 시험 연차에 따른 광의의 유전력(broad-sense heritability, h2)을 산출하였고(Toker, 2004), 산출식은 다음과 같다:
표현형 분산
유전력
(g:유전형, y:연차, r:반복, :유전형 분산, :유전형×연차 분산, :오차 분산)
또한 탈립 저항성으로 선발된 6계통 및 모·부본 3품종 간 생육특성 비교를 위하여 던컨의 다중 검정(Duncan’s multiple range test)을 수행하였다. 모든 통계 분석은 R program version 3.6.0 (The R Core Team, free software)을 이용하였다.
결과 및 고찰
건조 시간에 따른 내탈립성 변이
건조 시간별 실내 내탈립성 검정 결과 저항성 품종인 ‘대원콩’은 3년간 건조 후 모든 건조 시간에 개열한 협의 비율이 0%로 저항성을 나타내는 반면, 부본인 ‘새올’은 모든 건조 시간에 개열한 협의 비율이 100%로 감수성을 나타내었다. ‘다원콩’은 24, 48시간에 중도 저항성(30, 40%)을 나타내었으나, 72시간 후 3년 평균 개열한 협의 비율이 80%로 감수성을 나타내었다(Fig. 1). 건조기에서 24, 48, 72시간 후 개열한 협의 비율이 0%인 저항성 계통은 ‘대원콩’ב다원콩’ 조합에서 각각 57, 39, 26계통이었으며, ‘대원콩’ב새올’ 조합에서 각각 52, 35, 21계통이었다(Table 1). ‘다원콩’ 보다 ‘새올’이 초기 시간대에 더 탈립에 약한 특성을 나타내었고(Fig. 1), 각 집단 간 건조 시간별 탈립률을 비교하였을 때 ‘대원콩’ב새올’ 집단이 초기 시간대(24, 48시간)에 탈립률이 더 높은 것으로 나타나 탈립에 약한 것으로 판단되었다. 하지만 건조 72시간 후의 탈립률은 두 집단에서 통계적으로 유의한 차이를 나타내지 않았다(Table 2). 건조 72시간 후의 두 집단에서 탈립률에 유의한 차이가 없었다는 점과 건조 시간이 길어질수록 저항성인 계통과 감수성인 계통의 차이가 뚜렷하게 된다는 점을 감안하여 건조 72시간 후 개열한 협의 비율이 0%인 계통을 지표로 저항성인 계통을 선발할 수 있었다(Table 1).
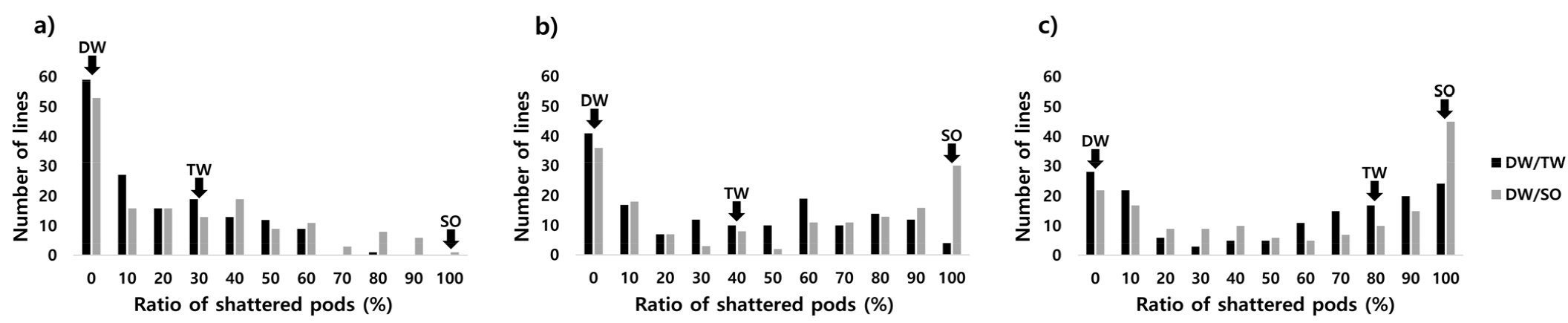
Table 1. Comparison of the ratio of average pod shattering potential of two populations in the pod drying test over three years.
Table 2. Comparison of the ratio of average pod shattering between two populations at different pod drying periods over three years.
| Combinationz | 24 h | 48 h | 72 h |
| D×T | 16.5 | 35.7 | 49.1 |
| D×S | 25 | 47.8 | 53.8 |
| t-valuey | -3.25** | -2.99** | -1.09ns |
y**denotes significantly different between the populations at p<0.01 by student t-test, ns denotes there was no significant difference
연차별 내탈립성 변이
시험 연차별 건조 72시간 후 개열한 협의 비율 또한 저항성 계통과 감수성 계통이 뚜렷하게 구분되는 경향을 나타내었다. 이러한 경향은 연차 간 비슷한 양상을 나타내었다(Fig. 2). 저항성 품종인 ‘대원콩’은 3년간 건조 72시간 후 개열한 협의 비율이 0%로 탈립에 매우 강한 특성을 나타내었고, 감수성인 ‘다원콩’은 개열한 협의 비율이 3년 평균 80%, ‘새올’은 100%로 탈립에 약한 특성을 나타내었다(Fig. 2). 각 연차별로 저항성 계통은 ‘대원콩’ב다원콩’ 조합에서 ’16년, ’17년, ’18년 각각 57, 50, 36계통이었고, ‘대원콩’ב새올’ 조합에서 42, 53, 24계통이었다(Fig. 2).
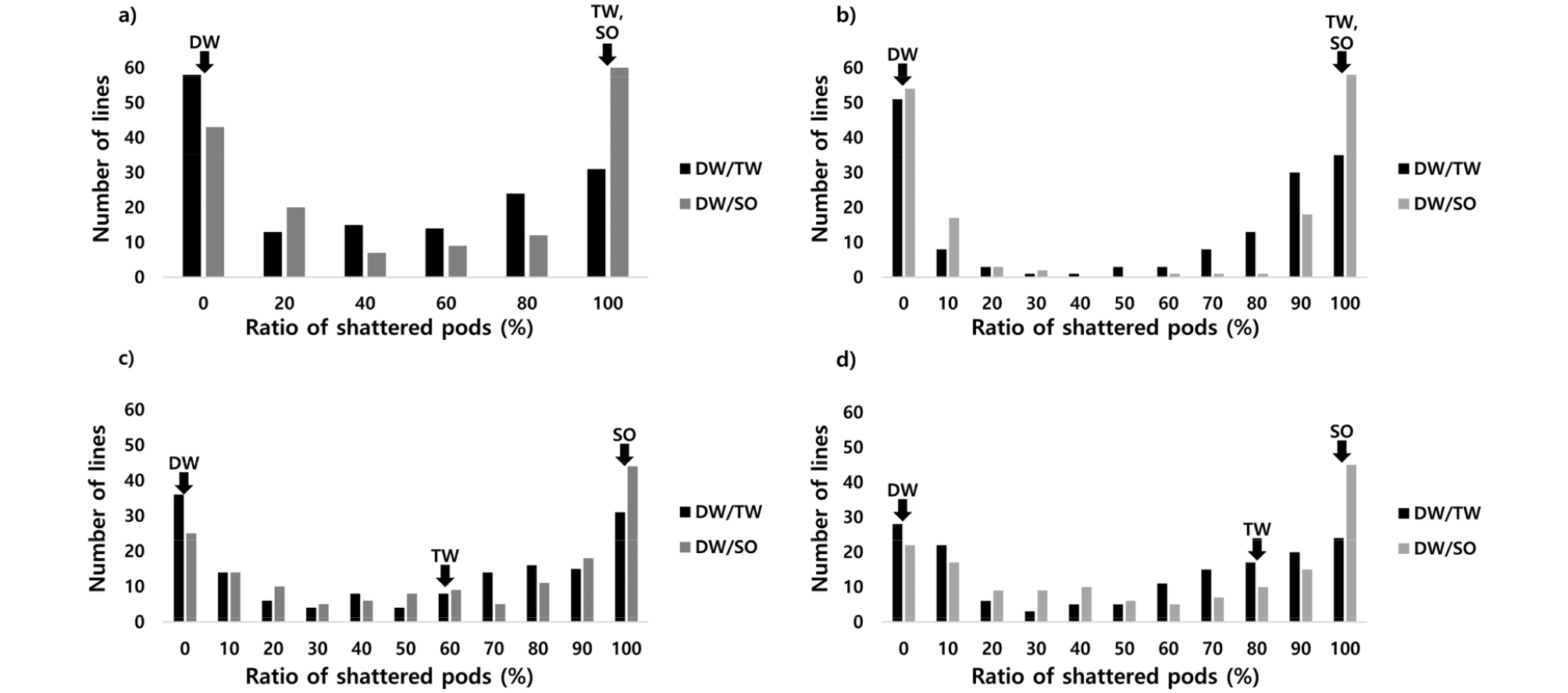
‘대원콩’을 모본으로 한 두개의 RIL집단에서 내탈립성 변이는 저항성 계통과 감수성 계통이 뚜렷하게 구분되는 빈도분포를 나타내어 질적형질에 가까운 형태를 나타내었다(Figs. 1 and 2). 따라서 ‘대원콩’의 내탈립성은 소수의 유전자에 의해 조절되고 이중에서도 탈립성에 강하게 관여하는 주동유전자가 존재한다는 것을 추측해 볼 수 있고, 이는 연차 간에 안정적인으로 것으로 판단된다. 이러한 결과는 16번 염색체상에 주동유전자가 존재한다는 기존 보고(Funatsuki et al. 2014, Lee et al., 2017)와 일치하였다.
처리 시간에 따른 효과와 계통 간 내탈립성 변이
본 연구에서 나타난 내탈립성 변이는 기존 보고(Kang et al., 2008) 보다 저항성과 감수성 계통 간의 탈립성 차이가 더욱 분명하게 구분되었고, 건조 시간이 증가할수록 이러한 경향은 더욱 뚜렷해졌다(Figs. 1 and 2). 건조 시간별, 연차별 내탈립성 변이에 대한 상관 관계 및 내탈립성에 영향을 미친 요인에 대한 유의성을 평가하기위하여 상관 분석(Figs. 3 and 4) 및 분산 분석(Table 3)을 수행하였다.
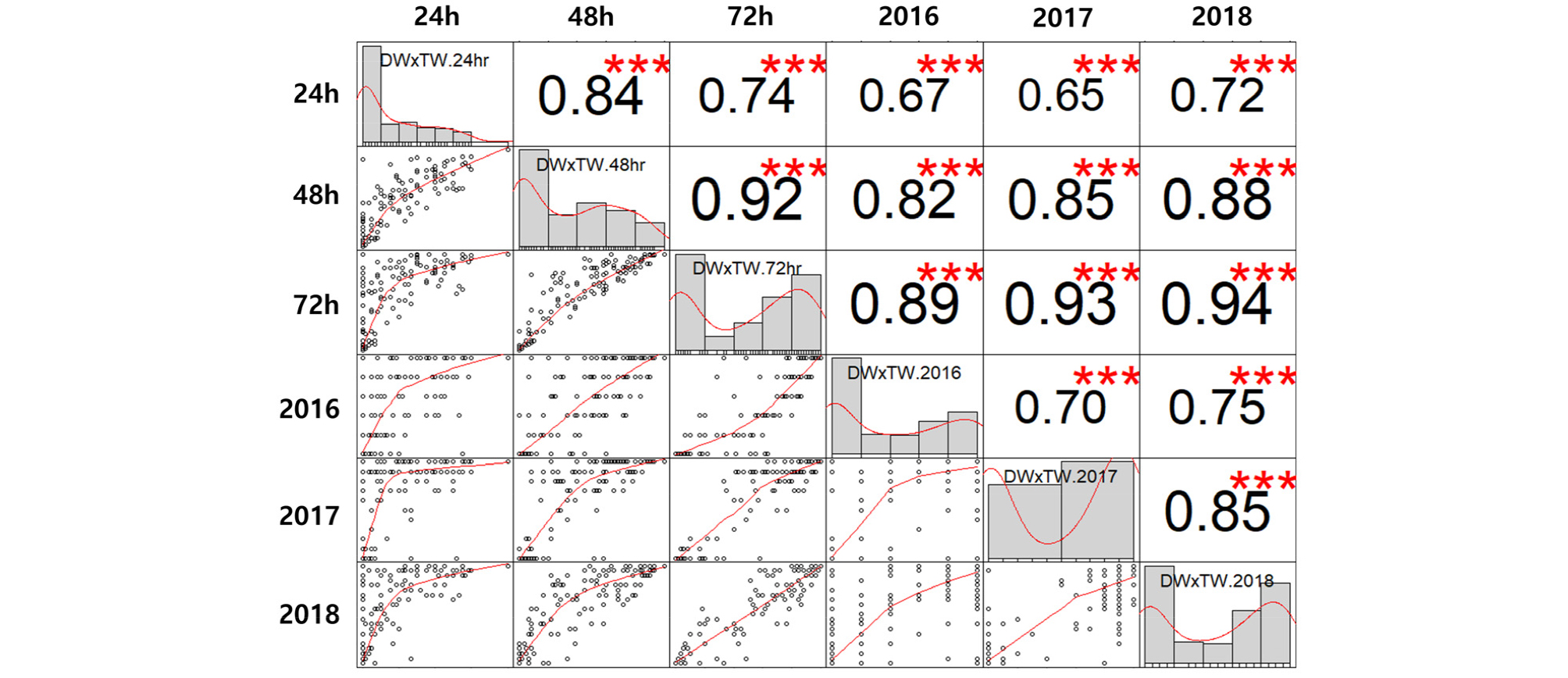
Fig. 3.
Correlation of pod shattering ratio between the pod drying periods (24 h, 48 h and 72 h, average pod shattering ratio over 3 years) and experimental years (2016, 2017 and 2018, pod shattering ratio drying after 72h) in RIL population derived from ‘Daewonkong’ (DW)בTawonkong’ (TW). ***denotes statistically significant values (p<0.001).
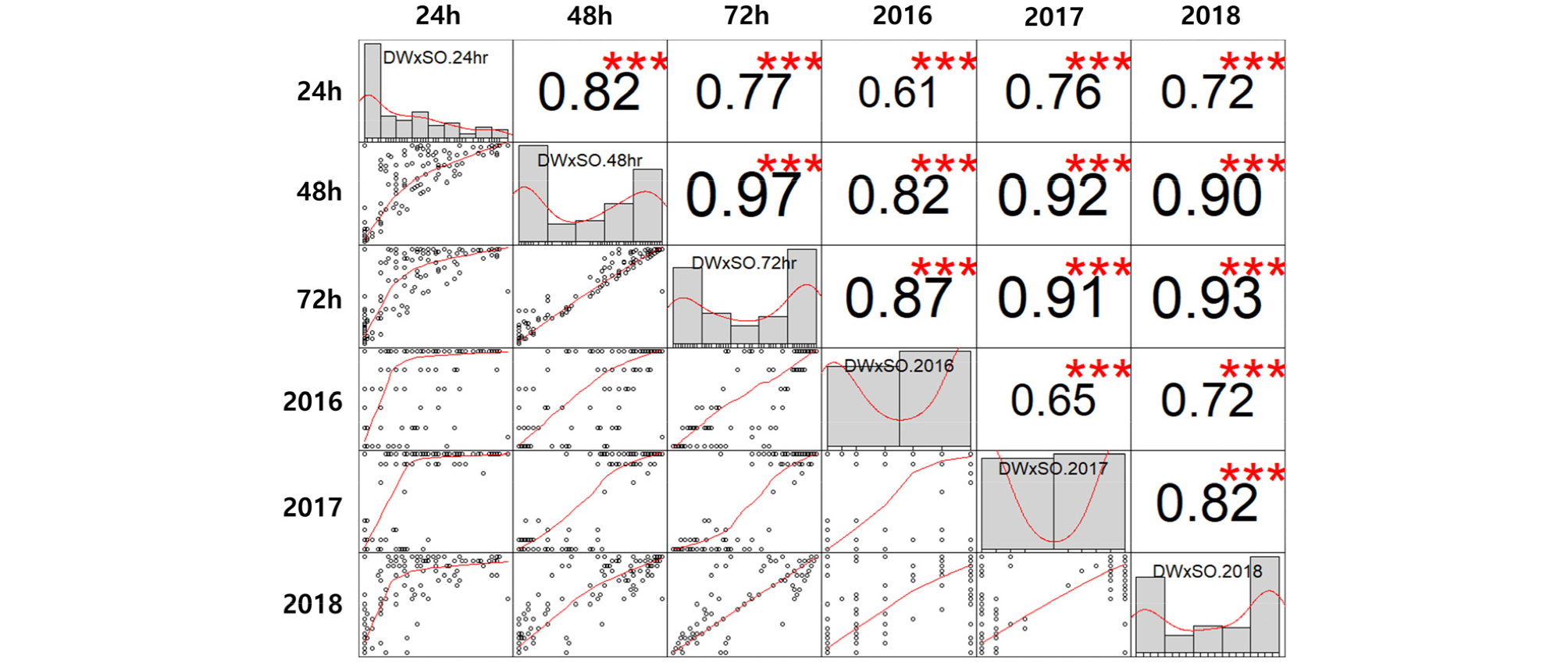
Fig. 4.
Correlation of pod shattering ratio between the pod drying periods (24 h, 48 h and 72 h, average pod shattering ratio over 3 years) and experimental years (2016, 2017 and 2018, pod shattering ratio drying after 72 h) in RIL population derived from ‘Daewonkong’ (DW)בSaeolkong’ (SO). ***denotes statistically significant values (p<0.001).
Table 3. Analysis of variance and broad-sense heritability for average pod shattering ratio over three years.
건조 시간별 상관분석 결과 두 집단에서 모두 건조 48시간 후의 내탈립성 변이와 72시간 후의 내탈립성 변이 간의 상관계수가 가장 높았고(r=0.92, p<0.001, r=0.97, p< 0.001), 다음으로 건조 24시간 후의 내탈립성 변이와 48시간 내탈립성 변이 간의 상관계수가 높았다(r=0.84, p<0.001, r=0.82, p<0.001). 또한 건조 24시간 후의 탈립성 변이와 72시간 후의 탈립성 변이 간의 상관도 높은 것(r=0.74, p<0.001, r=0.77, p<0.001)으로 나타났다(Figs. 3&4). 연차별 내탈립성 변이는 각 연차별로 유사한 형태의 빈도 분포를 나타내었고(Fig. 2), 상관 분석 결과 두 집단에서 탈립률에 대한 연차 간 상관계수는 r=0.65***에서 r= 0.85***로 높은 상관계수를 나타내었다(Figs. 3&4). 특히 ’16, ’17년도의 상관계수(r=0.70, p<0.001, r=0.65, p<0.001) 보다 ’17, ’18년도 간(r=0.85, p<0.001, r=0.82, p <0.001)의 상관계수가 높은 것을 확인할 수 있는데, 이는 연차가 지날수록 각 계통의 유전형이 고정되어 표현형이 더욱 안정적으로 나타나는 것으로 추측해 볼 수 있다.
시험 계통 및 연차 간 분산 분석 결과, 계통 간(genotype)에는 통계적으로 유의한 차이(p<0.001)를 나타내어 계통 간에 내탈립성 차이가 있다는 것을 알 수 있었고, 시험 연차 간(year)에는 유의한 차이를 나타내지 않아 ‘대원콩’ 유래의 내탈립성 변이가 연차 간에 통계적으로 비슷하여 안정적인 것을 알 수 있었다. 또한 계통과 연차 간(genotype×year)에는 상호작용(p<0.001)을 나타내어 계통에 따라서 연차 간 변이가 달리 반응하는 경우도 있음을 알 수 있었다(Table 3). 3년간 내탈립성에 대한 유전력은 0.86으로 높은 것으로 나타나 연차 간의 환경 변이 보다는 유전에 의한 계통 간의 내탈립성 차이가 더 큰 것으로 판단되었다.
생육특성 조사 결과
’16-’17년 2년간 두 개의 RIL 집단에서 내탈립성 계통으로 선발된 71개의 유망계통에 대한 생육특성 빈도분석 결과 각 형질별로 정규분포 형태의 분포를 나타내었다(Fig. 5). 따라서 내탈립성에 대한 선발 자원들 간에도 주요 생육특성 간에 차이가 있을 수 있으며, 내탈립성을 가지면서도 개화기가 빠르거나 생육특성이 우수하여 수량성 증진에 유리한 형질들을 집적하고 있는 계통들을 선발할 수 있었다.
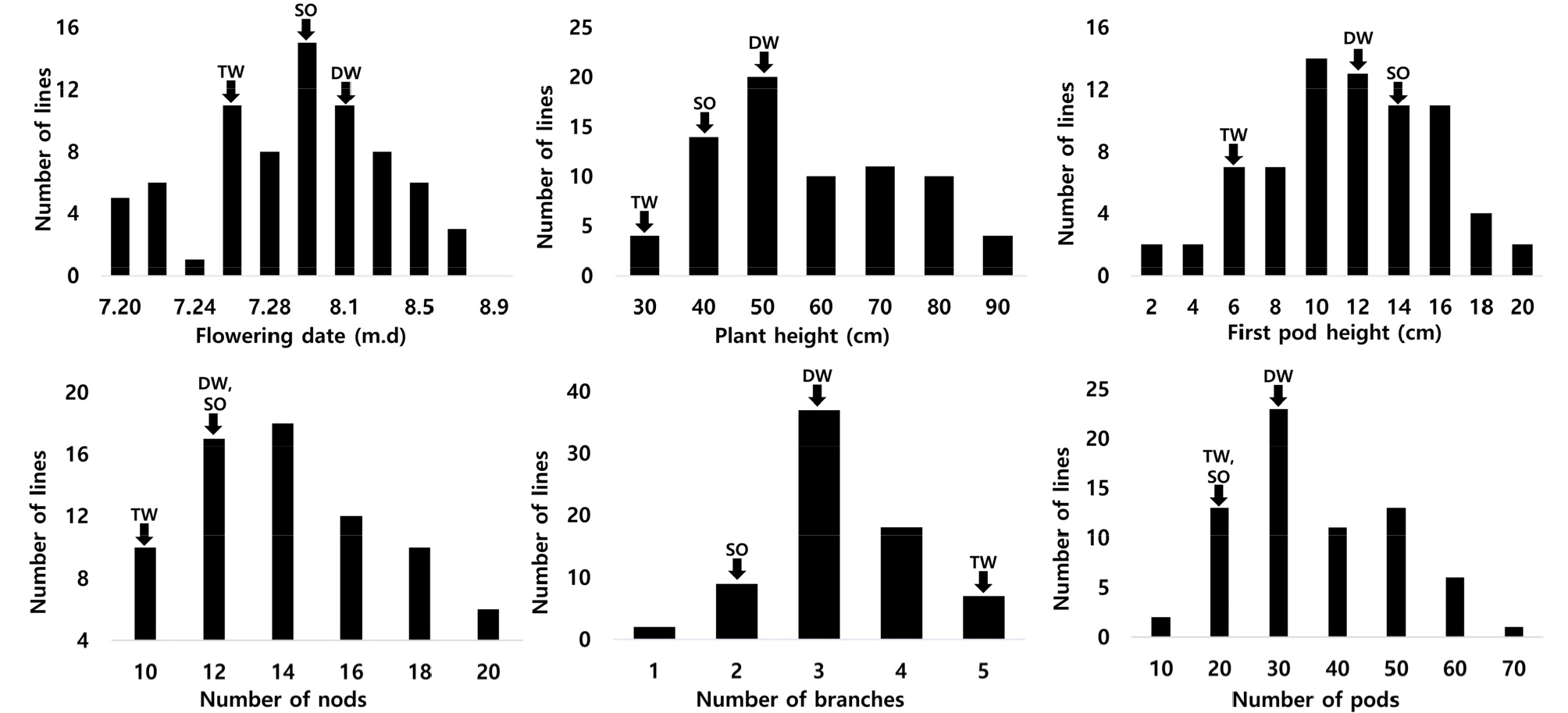
개화기는 ‘다원콩’이 7월 25일로 사용된 모·부본 중에 가장 빨랐고, ‘다원콩’ 보다 개화기가 빠른 계통은 12계통이었다. 모·부본 보다 경장 및 착협고가 높은 계통은 각각 38, 20계통이었으며, 모·부본 보다 마디수, 분지수, 협수가 많은 계통은 각각 40, 2, 45계통으로 조사되었다(Fig. 5). 이처럼 마디수, 분지수, 협수 등이 많은 계통은 ‘대원콩’ 수준의 내탈립성을 가지면서도 수량성이 증진될 것으로 생각된다.
내탈립성 유망계통 선발
’16-’18년 3년간 실내 내탈립성 검정 결과 건조기에서 72시간 후 개열한 협의 개수가 ‘0’인 저항성 계통은 ‘대원콩’× ‘다원콩’ 조합에서 26계통, ‘대원콩’ב새올’ 조합에서 21계통이었으며(Table 1), 이 중에서 개화기 및 성숙기가 ‘새올’에 준하여 빠른 단기성 계통과 종피색이 유색인 유색콩 계통을 각각 선발한 후 생육조사 결과와 종합하여 최종적으로 총 6계통을 Table 4와 같이 선발하였다. 선발된 6개 계통은 모본인 ‘대원콩’ 보다 성숙기가 7~15일 빨랐고, ‘18R49- 038’, ‘18R49-094’, ‘18R50-058’, ‘18R50-098’ 등 4계통은 경장 및 착협고가 높고 마디수가 많은 특성을 나타내었고, ‘18R49-035’, ‘18R50-059’ 등 2계통은 경장 및 마디수가 적지만 분지수가 많은 특성을 나타내었다. 또한 6개 계통 모두 협수가 모·부본 보다 많아 수량성이 증진될 것으로 판단되었다(Table 3). 이중에서도 ‘18R49-035’ 계통은 종피색이 갈색이면서도 개화기 및 성숙기가 빨라 내탈립성을 가진 유색 단기성 계통으로서 유망한 것으로 평가되었다.
Table 4. Average values of agronomic traits of the tolerant lines for pod shattering over three years.
yMean value of the ratio of shattered pods tested after 72 h of drying for three years
xMean values followed by the same letters in a column were not significantly different by Duncan's multiple range test
wF-values from analysis of variance. ***denotes significant difference (p<0.001)
적 요
콩의 내탈립성은 성숙기 수량 손실의 주요인이며 기계수확 시 장애요인으로서 개량되어야 할 불량형질로 간주된다. 국립식량과학원에서 개발된 ‘대원콩’은 성숙기 탈립에 매우 강한 특성을 나타내어 현재까지도 가장 널리 보급되어 있는 콩 품종이다. 향후 육종 프로그램에서는 ‘대원콩’ 수준의 내탈립성을 가지면서도 수량성, 품질 등이 개선된 품종을 개발하는 것을 목표로 하고 있다. 따라서 본 연구는 ‘대원콩’ 유래의 내탈립성에 대한 변이 양상을 조사하고, ‘대원콩’ 수준의 유색콩 및 단기성 콩을 선발하기 위해 수행되었다. ‘대원콩’을 모본으로 하고, 감수성인 검정콩 품종 ‘다원콩’과 단기성 콩 품종인 ‘새올’을 각각 부본으로 하는 두개의 고세대 RIL 집단을 활용하여 ’16년부터 ’18년 3년간 실내 내탈립성 검정을 수행하였다. 그 결과 건조 시간별, 연차별 내탈립성 변이가 두 집단에서 모두 저항성 계통과 감수성 계통이 뚜렷하게 구분되는 비슷한 분포를 나타내었고, 이에 따라 ‘대원콩’의 내탈립성은 기존 보고와 같이 소수의 유전자에 의해 조절되고 특히 가장 강하게 관여하는 유전자가 존재함을 알 수 있었다. 또한 ‘대원콩’ 유래의 내탈립성은 연차 간에 안정적인 형질이었으며, 건조 시간이 오래될수록 안정적이었다. 따라서 건조 72시간 후의 터진 협의 비율을 지표로 성숙기 탈립에 저항성인 총 47계통을 선발할 수 있었으며, 생육조사 결과와 종합하여 최종적으로 내탈립 유색콩 및 단기성콩 6계통을 선발할 수 있었다. 선발된 6개 계통은 ‘대원콩’ 수준의 내탈립성을 가지면서도 성숙기가 빠르고, 경장, 마디수, 협수 등이 높아 추후 육종프로그램에서 내탈립성인 유색콩, 단기성콩 품종 개발의 모본으로서 유용하게 활용될 것으로 기대된다.
