

서 론
재료 및 방법
재배 방법
기상 분석
예측모델을 통한 토양수분 분석
생육 및 수량조사
지방 및 단백질 분석
통계분석
결과 및 고찰
각 연도별 생육 시기 및 기상
토양수분 모형(AFKAE0.5)을 통한 토양 수분함량 예측
연도별 특징에 따른 콩의 생육 및 수량
각 품종에 따른 연도별 품질 비교
적 요
서 론
콩은 우리나라에서 오랜 기간 동안 재배되어 오면서 각종 장류, 두부, 콩나물 등 여러가지 식품으로 이용되고 있고 우리나라를 포함한 각 나라의 전통식품으로 이용되고 있다(Coward et al., 1993; Messina & Messina, 2010). 또한 콩에는 다량의 단백질과 지방을 함유하고 있고 칼슘, 인산 등 다양한 무기영양소가 풍부하여 식생활에 있어 많은 영양을 공급해주는 중요한 작물 중 하나이다. 그러나 기후변화로 인해 지구온난화와 이상 기상이 발생하여 콩을 포함한 모든 작물의 수량이 감소하는 등 작물 피해가 전 세계적으로 증가하고 있다. 또한 온난화로 인해 지속적으로 평균 기온이 높아지거나 이상 기상 발생 빈도가 증가할 것으로 전망하고 있어(Im et al., 2015), 이러한 환경변화에 대응하기 위해 다양한 연구가 진행되고 있다.
기온 상승에 따른 콩의 생육 및 수량 관련 연구는 오랜 기간 동안 진행되고 있다. Thanacharoenchanaphas & Rugchati (2011)는 생식생장기에 환경이 좋지 않을 경우 콩의 수량이 감소한다고 하였고, Reddy & Kakani (2007)는 생식생장기가 영양생장기보다 온도에 더 민감하다고 보고하였다. 또한 Egli & Wardlaw (1980)은 등숙기간 중 주야간 온도가 33/28°C일 경우 24/19°C보다 종실 수량이 약 22% 감소한다고 하였다. Puteh et al. (2013)은 콩의 생육 및 수량 감소는 고온 노출 기간과 생식생장기에 의해 영향을 받으며, 개화시부터 종실비대시동안 고온에 오래 노출될 경우 수량에 큰 영향을 준다고 보고하였다. 또한 생식생장기 중 개화기의 고온은 꽃의 불임을 증가시킨다고 하였고(Djanaguiraman et al., 2013) 적정 온도에 비해 착협률 및 착협수를 감소시키며(Allen et al., 2018), 종실비대시 기간의 고온은 종실중을 감소시킨다고 하였다(Thomas et al., 2010; Tacarindua et al., 2013; Choi et al., 2016).
또한 기온상승에 따른 콩의 지방과 단백질 함량에 대한 연구도 활발히 진행되고 있다. Lee et al. (2018)은 종실비대기에서 성숙기까지 평균기온이 25°C 이상일 경우 대원콩의 지방 및 단백질의 함량이 줄어들고, 풍산나물콩은 평균기온이 21°C 이상 올라갔을 때 지방 함량이 감소하고 23°C 이상 올라갈 경우 단백질 함량이 감소한다고 보고하였다. Song et al. (2016)은 낮 기온이 19.7°C일 경우 지방 함량과 평균 온도 사이에 상관관계가 존재한다고 하였고 단백질 함량은 적산온도 15°C 이상의 일교차에서 비례하지만 일조 시간과 반비례한다고 보고하였다. 콩과 수분스트레스 관계에서 다양한 연구결과가 있는데 Kumar et al. (2006)와 Rotundo & Westgate (2018)는 토양 수분이 부족할 경우 단백질 함량이 증가한다고 보고한 반면에 Specht et al. (2001)와 Boydak et al. (2002), Carrera et al. (2009)은 토양수분 부족이 단백질 함량을 감소시킨다고 보고하였다.
해외에서는 온도 및 수분 스트레스에 의한 콩의 연구가 다양하지만 국내에서는 이에 대한 연구가 부족한 실정이다. 본 연구는 기상이 서로 다른 연도에 따른 콩의 생육 및 수량구성요소를 조사하고 지방 및 단백질 함량을 분석하여 기상에 따른 수량 및 품질 변화에 대한 기초자료로 활용하고자 수행하였다.
재료 및 방법
재배 방법
본 시험은 전라북도 완주군 이서면 국립식량과학원 시험포장에서 2017년부터 2018년까지 수행하였다. 품종은 장류콩인 대원, 대풍2호와 나물콩인 풍산나물콩을 사용하였고, 파종 날짜는 2017년과 2018년 각각 6월 20일에 파종하였다. 재식 거리는 대원과 대풍2호의 경우 70×15 cm, 풍산나물콩은 70×10 cm로 설정하여 농촌진흥청 표준영농교본에 준하였으며(RDA, 2015), 시비량은 30-30-34 kg·ha-1 (N-P-K)이었다.
기상 분석
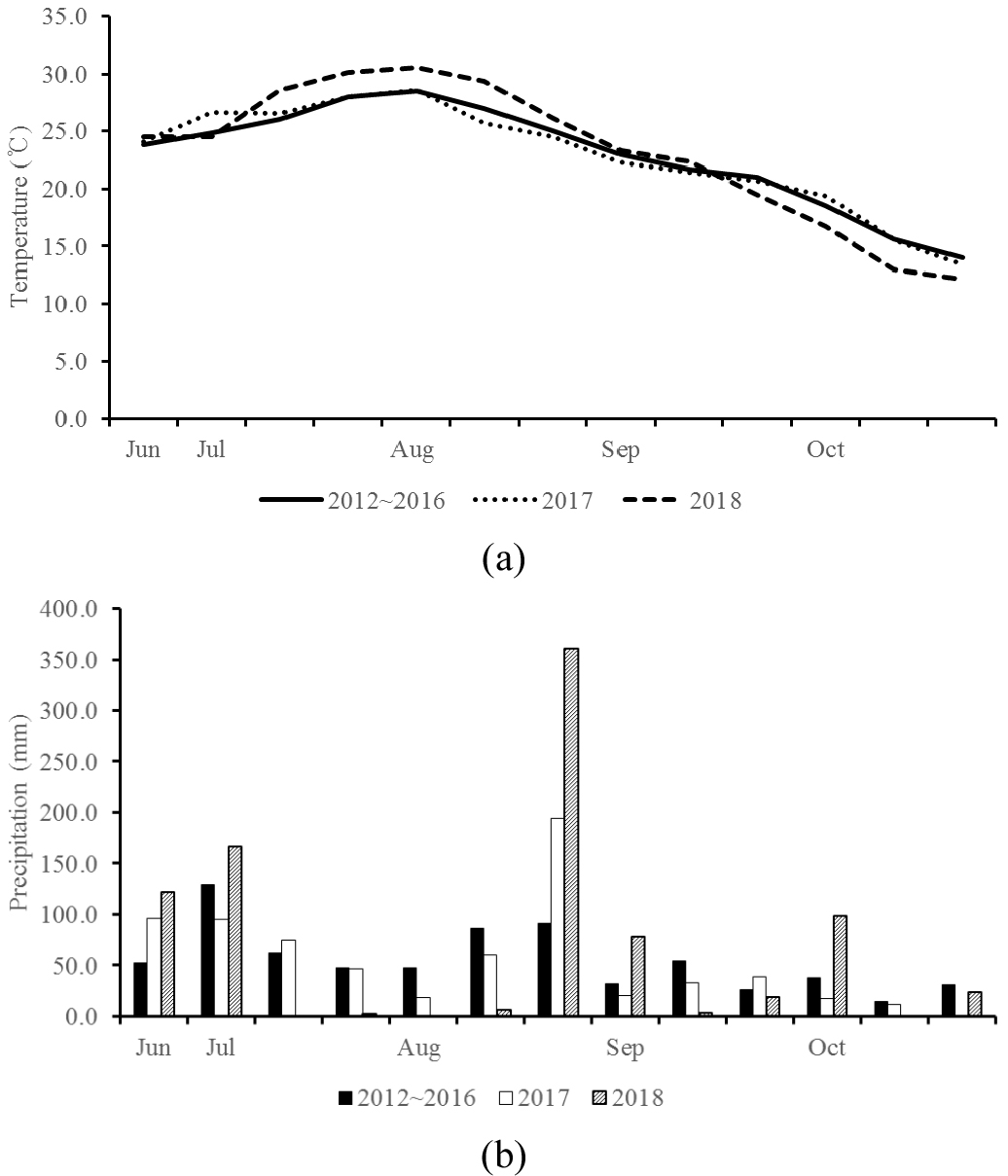
재배기간 동안의 기상 자료와 평년 기상은 2012년부터 2016년까지의 자료는 전주지역 기상청 기상 자료로 분석하였다. Fig. 1은 2017년과 2018년의 평균온도(a)와 강수량(b)을 평년 기상과 비교하여 나타낸 것이다.
예측모델을 통한 토양수분 분석
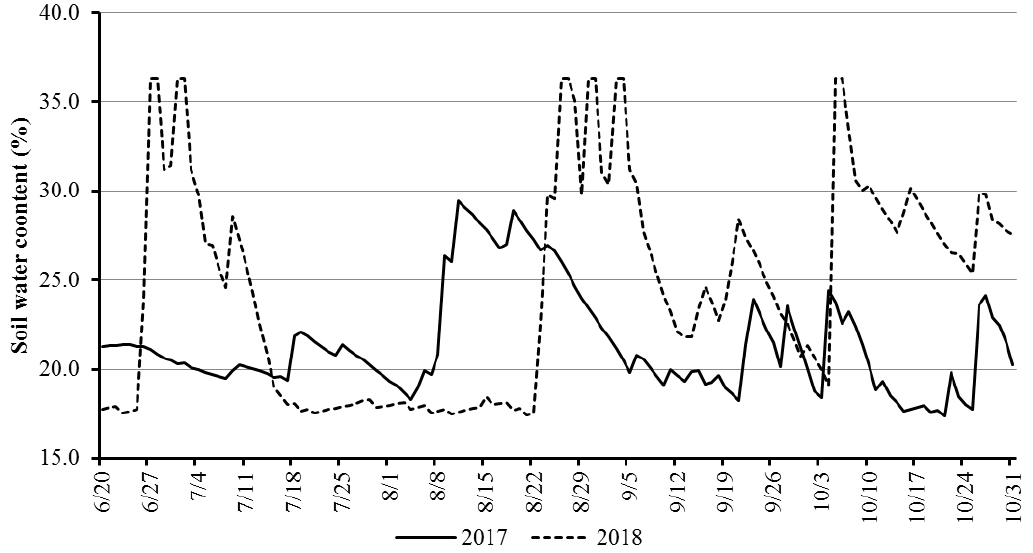
2017년과 2018년에 재배된 기간 동안 시험포장에서 토층 300 mm 이하의 수분함량을 알아보기 위해 토양수분 예측 모형인 AFKAE0.5 (Seo et al., 2012)를 사용하여 토양수분함량을 예측한 후 각 연도별 수분함량 예측 결과를 분석하였다.
생육 및 수량조사
콩 수량 구성 요소 조사는 10월 20일에 품종 당 식물체 20주를 3지점 채취하였다. 조사방법은 농업과학기술 조사분석기준(RDA, 2012)에 준하여 경장, 경태, 마디수, 분지수, 개체당 착협수, 개체당 립수 및 100립중, 수량을 조사하였다. 종실 크기는 건조시킨 300개의 종실을 test sieve를 이용하여 4 mm부터 8 mm까지 나누어 그 비율을 퍼센트로 나타내었다.
지방 및 단백질 분석
연차와 품종에 따른 조지방 분석은 조지방 자동 추출 장치(Foss, Soxtec8000, Hilleroed, Denmark)를 이용하여 마쇄한 종실 3 g을 넣은 원통 여지(Thimble filter)에 Ether를 넣어 추출한 후 105°C 항온기에 1시간 30분동안 Ether를 제거하였다. 건조시킨 원통 여지는 데시게이터에 옮겨 30분 냉각한 후 무게를 측정하였다. 단백질 분석은 Dumas법(Fred & Watts, 1993)으로, 마쇄한 종실을 0.1 g 칭량하여 원소분석기(Leco, FP628, U.S)로 측정한 값을 단백질 환산 계수 6.25를 곱하여 나타내었다.
통계분석
연구결과에 따른 통계 분석은 RStudio ver. 1.2.1335를 활용하여 분산분석을 통해 p < 0.05수준에서 유의성을 검정하였다.
결과 및 고찰
각 연도별 생육 시기 및 기상
2017년과 2018년의 품종별 생육 시기는 Table 1과 같다. 개화기는 대풍2호가 연도에 관계없이 가장 빠른 경향을 보였고, 개화기가 빨랐기 때문에 착협시(R3), 종실비대시(R5) 등 생육 시기가 빨라진 것으로 판단된다. 반면 풍산나물콩은 대원, 대풍2호에 비해 각 생육 시기가 연도별로 큰 차이를 보이지 않았다.
Table 1.
Growth period of each cultivar in 2017 and 2018.
VE: Vegetative stage emergence, R1: One flower at any node, R3: Pod 0.5cm long at one of the four uppermost nodes with a completely unrolled leaf, R5: Seed beginning to develop at one of the four uppermost nodes with a completely unrolled leaf, R7: Pod yellowing; 50% of leaves yellow. Physiological maturity
Table 2는 두 연도의 평균 기온과 강수량을 콩의 생육시기별로 나타낸 것이다. 2017년과 2018년의 기온을 살펴보면 출현기~개화시(VE~R1)의 기온은 대원과 대풍의 경우 차이가 거의 없었고 풍산나물콩은 0.9°C 차이가 났는데 2018년 7월 하순의 평균기온이 2017년보다 높은 것이라 판단된다. 개화시~착협시(R1~R3)의 경우 2018년의 기온이 2017년보다 모든 품종에서 약 2°C 이상 높았고 착협시~종실비대시(R3~R5)에서 2018년 대원과 대풍2호의 평균기온이 2017년보다 각각 3.4, 3.6°C 높았으나 풍산나물콩은 1.9°C 높아 다른 품종에 비해 연도별 온도차이가 적었다. 종실비대시~성숙기(R5~R7)에서 대원과 대풍2호의 2018년 평균기온은 2017년보다 각각 1.3, 1.4°C 높았으나 풍산나물콩은 두 연도별 차이가 거의 없었다. 두 연도를 기온을 비교해보았을 때 개화시 이후 고온으로 인한 스트레스가 일어났을 것이라 예상되고 품종 중에서 대원과 대풍2호가 풍산나물콩에 비해 스트레스를 많이 받았을 것이라 생각된다. 2017년과 2018년의 강수량을 생육시기별로 살펴보면 출현기~개화시의 경우 두 연도에서 200 mm 후반대로 나타났고 2017년 풍산나물콩에서 300 mm대로 나타났다. 개화시~착협시의 강수량은 2017년의 경우 모든 품종에서 30 mm 이상 내렸지만 2018년의 경우에는 모든 품종에서 2 mm 미만으로 내려 강수량이 2017년보다 매우 적었다. R3~R5의 강수량은 2017년에 모든 품종에 200 mm 이상의 비가 내렸으나 2018년에는 대원이 57.9 mm, 대풍2호가 7.3 mm 정도 내려 2017년에 비해 적게 내렸다. 풍산나물콩은 368.6 mm가 내렸는데 이는 8월 하순에 강수량이 많았기 때문이다. Allen et al. (2018)은 콩을 고온에서 재배했을 때 적정 온도에 비해 착협률과 착협수가 감소한다고 보고하였고 Van Doren & Reicosky (1987)은 생육기간 중 개화시부터 종실비대시까지 수분이 가장 많이 필요하다고 보고하였다. 이런 요인으로 인하여 2018년에는 콩의 생육시기 중 개화시부터 종실비대시까지 기간동안 기온상승과 강수량 부족으로 인해 콩의 생육과 수량에 영향을 미쳤을 것으로 생각된다.
Table 2.
Average temperature and total precipitation of cultivar during the growth stage in 2017 and 2018.
VE: Vegetative stage emergence, R1: One flower at any node, R3: Pod 0.5cm long at one of the four uppermost nodes with a completely unrolled leaf, R5: Seed beginning to develop at one of the four uppermost nodes with a completely unrolled leaf, R7: Pod yellowing; 50% of leaves yellow. Physiological maturity
토양수분 모형(AFKAE0.5)을 통한 토양 수분함량 예측
토양수분 모형을 이용한 각 연도의 일별 수분함량 예측 그래프는 Fig. 2와 같다. 2017년의 수분함량 예측 그래프를 보면 대원과 대풍2호는 착협시, 풍산나물콩은 개화기에 포함되는 기간에 수분함량이 부족했지만 8월 8일부터 수분함량이 증가하는 것으로 예측되었다. 반면 2018년의 경우 대원과 대풍2호는 개화 이전부터 종실비대시까지, 풍산나물콩은 협이 생성되는 기간인 7월 중순부터 8월 중순까지 지속적으로 한발이 예측되었다. Doss et al. (1974)는 개화 초기에 수분이 부족할 경우 콩의 수량이 약 30% 감소하고 개화기부터 성숙기까지 수분이 부족할 경우 약 50% 감소한다고 하였고, Kim & Koh (1997)은 콩의 생육 시기 중 착협시에 한발이 생길 경우 다른 생육 시기에 비해 수량 피해가 크다고 보고하였다. 이와 같은 결과로 인해 한발에 의한 콩의 생육 및 수량 피해는 2018년에 있을 것이라 예상되었다.
연도별 특징에 따른 콩의 생육 및 수량
Table 3은 연도에 따른 각 품종별 생육 상태를 나타낸 것이다. 2017년의 경장은 모든 품종에서 2018년보다 컸고 직경 또한 2017년이 2018년보다 모든 품종에서 굵었다. 분지수는 2017년이 대원과 풍산나물콩에서 2018년보다 많았고 대풍2호의 경우 2018년이 2017년보다 0.5개 더 많았지만 다른 품종에 비해 큰 차이를 나타내지 않았다. 마디수는 경장, 직경과 마찬가지로 모든 품종에서 2017년이 2018년보다 많은 경향을 나타내었다 Park et al. (2010)은 고온에 황금콩과 풍산나물콩을 고온 환경에서 재배했을 때 대조구에 비해 생육이 증가했다고 보고했는데, 2018년의 평균온도가 2017년보다 높았지만 2018년의 생육이 2017년보다 좋지 않은 이유는 수분 부족으로 인해 생육에 안좋은 영향을 준 것으로 생각된다. 연도에 따른 품종별 수량구성요소는 Table 4와 같다. 식물체당 착협수를 보면 2017년이 모든 품종에서 2018년보다 많았다. 연도에 살펴보면 2017년의 립수가 2018년보다 모든 품종에서 많았고, 품종별로 살펴보았을 때 풍산나물콩 > 대풍2호 > 대원 순으로 많았다. 100립중은 착협수, 립수와 마찬가지로 모든 품종에서 2017년의 100립중이 2018년보다 컸으며, 품종별로 비교했을 때 대원의 100립중이 다른 품종보다 연도별 차이가 가장 컸다. 수량은 2017년의 수량이 2017년보다 많았고 세 품종을 연도별로 비교해본 결과 대원의 수량이 다른 2품종에 비해 수량이 가장 낮았다. 이는 온도가 높아질수록 개체당 협수가 감소한다고 보고한 Puteh et al. (2013)의 결과와 일치하였으며, 개화시부터 종실비대시 이전까지의 수분 스트레스가 다른 생육 기간보다 수량에 민감하다는 연구결과가 있어(Boyer, 1983; Westgate & Peterson, 1993), 고온과 한발로 인해 콩 수량에 영향을 줬을 것이라 판단된다.
Table 3.
Growth status of each cultivar during harvest period in 2017 and 2018.
Table 4.
Yield component of each cultivar during harvest period in 2017 and 2018.
Table 5는 연도별 각 품종의 종실 크기를 나타낸 것이다. 대원의 연도별 종실 크기를 보면 2017년의 경우 6.7 mm 이상 8 mm 미만의 직경 비율이 가장 높았지만 2018년의 경우 6.7 mm 이상 8 mm 미만 직경의 비율이 55.5%이었으며, 전체적으로 종실 크기가 2017년이 2018년보다 크다는 것을 알 수 있다. 대풍2호의 경우에는 5.6 mm 이상 8 mm 미만에서 2017년의 비율이 2018년보다 많았고 2018년의 종실 비율이 더 작아지는 경향을 나타내어 대원과 결과가 비슷하였다. 풍산나물콩은 2017년에서 5 mm 이상 5.6 mm 미만의 종실 크기 비율이 2018년보다 더 많았고 종실 크기가 상대적으로 작은 비율은 2018년이 더 많았다. 종실 크기를 연도별로 비교해 보았을 때, 직경이 큰 종실은 2017년이 2018년보다 많이 분포해 있고, 품종별로 비교했을 때 대원의 종실 비율이 다른 품종에 비해 작은 곳에 많았다. Liu (2004)는 적은 일사량, 한발 등과 같은 환경 스트레스가 종실 크기를 감소시킨다고 보고하였는데, 본 연구에서 2018년의 종실 크기가 2017년보다 작아진 원인은 콩의 생식생장기 중 개화시부터 종실비대시 이전까지 한발로 인하여 종실 크기가 작아진 것으로 판단된다.
Table 5.
Distribution of soybean seed for three cultivars in 2017 and 2018.
각 품종에 따른 연도별 품질 비교
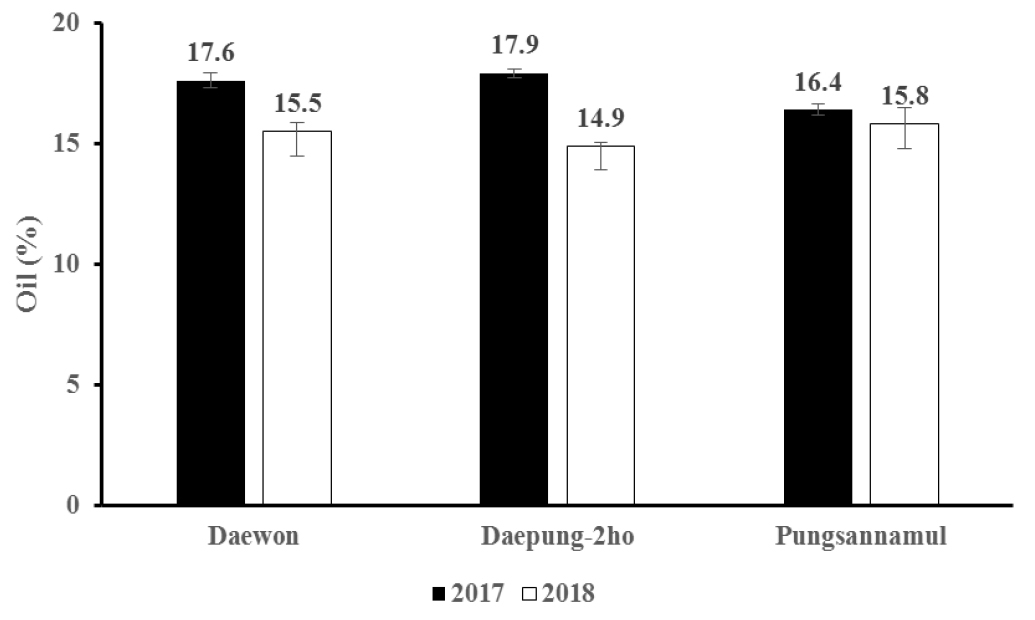
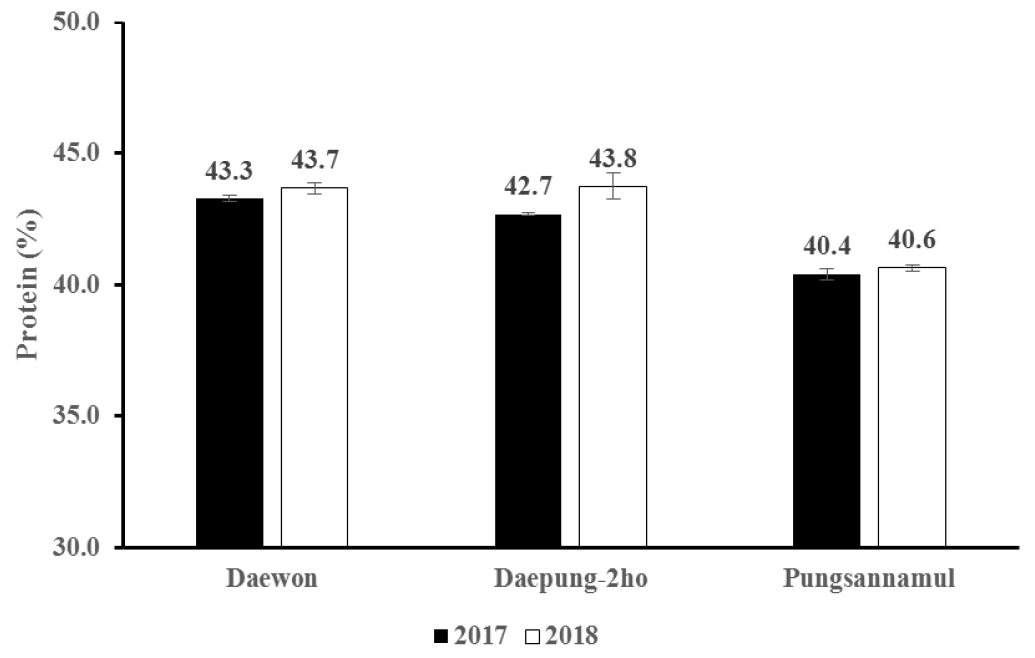
연도에 따른 품종별 지방 함량은 Fig. 3과 같다. 대원의 지방 함량은 2017년은 17.6%로 나왔고 2018년은 15.5%로 나타나 2017년의 지방함량이 2018년보다 많았다. 대풍2호의 지방 함량은 2017년의 경우 17.9%, 2018년의 지방 함량은 14.9%로 대원과 같은 현상을 보였지만 연도별 차이는 대원이 2.1%, 대풍2호가 3.0%로 대원보다 0.9% 더 컸으며 기온이 높거나 한발 스트레스가 일어났을 때 지방 함량이 낮아진다고 보고한 Dornbos & Mullen (1992)의 연구결과와 일치하였다. 그러나 풍산나물콩의 지방 함량은 2017년이 16.4%, 2018년이 15.8%으로 대원과 대풍2호에 비해 큰 차이를 보이지 않았다. 두 연도별 품종에 따른 단백질 함량은 Fig. 4와 같다. 대풍2호의 단백질 함량은 2017년이 42.7%, 2018년이 43.8%로 2018년이 2017년보다 더 많아 연도에 따른 차이를 나타내었다. 반면 2017년 대원의 단백질 함량은 2017년이 43.3%, 2018이 43.7%로 연도별 큰 차이를 나타내지 않았으며 풍산나물콩 또한 단백질 함량은 2017년이 40.4%, 2018년이 40.6%로 거의 비슷하였다. 결과적으로 두 연도별로 기온 및 강수량이 달랐지만 지방 및 단백질 함량에서 품종별로 다른 차이를 나타내었다.
적 요
본 연구는 2017년과 2018년의 기상을 가지고 연도별에 따른 대원, 대풍2호, 풍산나물콩의 생육 및 수량구성요소와 종자 크기, 지방, 단백질 함량을 분석하였다. 두 연도별 생육 시기를 보면 대원과 대풍2호에서 2018년이 2017년보다 생육 시기가 빠른 경향을 보였으나 풍산나물콩은 큰 차이가 없었다. 생육 기간 중 R1~R5에서 평균기온은 모든 품종에서 2018년이 2017년보다 높은 반면에 강수량은 풍산나물콩을 제외한 모든 품종에서 2018년이 2017년보다 적었다. 수분모형으로 각 연도의 일별 토양수분함량을 예측한 결과 2018년이 2017년보다 한발 기간이 길었고 개화시부터 종실비대기 이전까지 한발이 지속된 것으로 예측되었다. 종실 수량과 백립중은 2017년이 2018년보다 더 컸으며, 종실 크기는 2017년의 종실이 2018년보다 큰 경향을 나타내었다. 지방 함량은 모든 품종에서 2017년의 지방함량이 2018년보다 많았고 대풍2호, 대원이 연도별 기상에 따른 큰 차이를 보였다. 반면 단백질 함량은 2018년이 2017년보다 높은 경향을 보였지만 품종별 차이를 나타내었다. 결과적으로 2018년의 개화시부터 종실비대시 이전 기간이 2017년보다 평균기온이 높고 강수량이 적은 기상으로 인하여 콩의 수량, 지방 및 단백질 함량에 영향을 주었다. 이번 연구를 통하여 고온과 한발에 내성이 있음과 동시에 품질 감소가 저하되는 콩 품종 개발 및 재배 연구가 활발하게 이루어질 것이라 생각된다.
